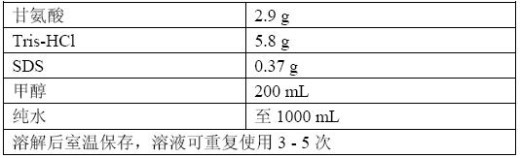
一、样品制备
1. 变性条件——SDS LB直接裂解
用常温或者高温预热过(预热更有利于阻止蛋白水解或者磷酸酶去磷酸化,但容易遗忘,LB久煮某些性质会改变)的undefinedSDSLB(或者略高1.undefined)直接加 到细胞或者组织上并煮样。通常6孔板细胞80%以上密度,需200ul,其他按平板面积比类推。通常条件下,SDSLB都是过量的(因此不一定要严格参考 SDS LB稀释比);如果SDSLB不够,样品核酸、蛋白浓度过高时,煮样后会发现tip吸不上来,非常粘、一砣一砣的,很容易堵住tip。
这时候常规做法有两 种,1.再煮5min。常规煮样时间3-5min,样品过浓时就煮10min;2.如果10min煮样后,仍然吸不起来,才适当增加SDSLB,继续煮; 也可以对样品进行超声。煮样时间若过长,蛋白会凝固,此时以失去继续WB的意义,请丢弃(判断标准:出现明显的蛋白沉淀和水分层)
此方法的缺点,SDS LB煮过的样品如果用来做IP,需要特殊方法,因此,这种制备方法制备的样品有使用局限。
非变性裂解法(不讨论诸如核抽提、亚细胞器分离等等了,其实类似)
裂解细胞请尽量冰上操作,减缓酶解作用。
俺前boss nature文章上的裂解液配方是:20mM Tris/HCl, pH7.6, 100mM NaCl, 20mM KCl,1.5mM MgCl2, 0.5% NP-40 and protease inhibitors (20μg/ml leupeptin,10μg/ml pepstatin A and 10 μg/mlaprotinin)(此裂解液可用于抽提核蛋白),其中蛋白酶抑制剂很复杂也很贵,其实用0.5mMPMSF替代这三种就好了;如果还要做磷酸化蛋白,加1mM Na3VO4(现加现用),10 mM NaF, 1 mM Na3VO4, 50 mMbeta-GP。
此方法的优点:NaCl浓度略低于生理状态,保证裂解细胞的方式较为温和,便于随后的IP等试验。其他Na盐浓度的细胞裂解液也很常见,诸如 0.15M(生理盐浓度)、0.3M、0.6M(高盐系列,裂解细胞时染色体会析出,细胞碎片和沉淀非常粘稠)及0.8-1.2M;0.3M及以下适合 IP试验,0.6M及以上适合纯化蛋白,尤其相互作用较强的复合体,要得到纯化的一些复合体的组成蛋白一般采用极端的高盐裂解细胞。
用于核蛋白的裂解的原因是0.5% NP-40,用于破坏核膜结构;可用到1%,但此时染色体会析出,沉淀粘稠。
2. 组织块裂解
组织块较大用匀浆的方法最合适,现在有不少转头很小的匀浆器。
当组织超微量的时候,比如50mg新鲜癌组织,我采用的方法是
1. 低温(剪)搅碎,成肉糜状。
2. 锡箔纸包裹好,放入少量液氮,快速敲击(控制力量,尽量避免弄破锡箔纸),进一步破碎;反复多次。
3. 用上述细胞裂解液回收。
分泌型蛋白富集:
用无血清培养基培养细胞,收集上清液用于WB检测;如果含量过低,需要用TCA沉淀富集。
理论上,从普通培养基中收集分泌蛋白,此时对细胞生长条件的影响最小,是最佳的实验条件;但是这存在隐忧。因为含血清的培养基中含有非常丰富的BSA(牛 血清白蛋白)之类的蛋白质,除非你进一步分离纯化,否则直接用这种样品去WB会有非常大的麻烦,尤其当你的蛋白在60-46kDa之间的时候;用“任何” 抗体去检测这一区间的蛋白,会有一片非常强的信号。
因为此区间蛋白(主要是BSA)浓度过于富集,会非特异性粘附“任意”抗体,从而最终被识别。同理,采 用SDS LB裂解细胞制备样品前,通常要用PBS洗涤,其目的之一是去除培养基中的BSA。
采用无血清培养基收集细胞上清除了改变细胞的生理状态外(可能激活未知信号途径,诸如AKT等),还会出现的问题是:
1. 即使采用了无血清收集上清,仍然有大量杂蛋白,但相对好很多。
2. 选用了上清,由于蛋白浓度太稀,通常要采用TCA沉淀富集,再重新溶解。
a. TCA沉淀对半定量操作要求非常高,因为很容易沉淀不充分或者离心操作丢失,尤其在离心过程,离心管的摆放非常讲究,要180度翻转离心两次。
b。重新溶解时,由于蛋白需要在一定的盐离子条件下才能重新溶解,你要摸索充分溶解的条件,相对麻烦。
实际上,如果你不是非常强调蛋白的分泌有什么生理功能,一般不建议检测上清,尤其半定量的WB实验检测不同样品间的差异。这里面有实验操作细节的麻烦——经验之谈,我曾经参与的cancer cell文章所研究的蛋白就是分泌型蛋白,但90%的数据是celllysis;偶尔用到上清,也只是作为富集纯化该蛋白的原料,为进一步分析做准备。
样品制备完,应立即低温保存(-20度短期(几天);-80度长期;例外,IP用样品应直接进行IP,避免冻融破坏蛋白质间弱的相互作用)。SDSLB煮沸过的样品冻融会存在另一个问题,SDS沉淀;SDS 4度就会沉淀,何况-80度。上样前应加热充分溶解,否则从加样口向下拖带(SDSLB不够,样品未充分溶解也会出现类似拖尾;上样过大也会)。
在样品制备过程中,另一个需要注意的问题是,从样品制备的起始阶段就要注意定量问题;WB本身系统误差有20%,太细微的差别经常忽略不计。
细胞样品可以 先计数再接种,短时间内即使细胞生长有差异也不会对WB结果影响太多。组织样品,从肿瘤组织的大小(称重)开始,裂解液的体积都是精确定量的,操作过程也 尽量避免蛋白损失。有人会说可以电泳前蛋白定量,我将下一节讨论蛋白定量的不科学性,以及裂解液成分对定量的干扰。
蛋白电泳:
电泳胶的制备、配方、缓冲液配方,请参考分子克隆。经验之谈,8%胶最底边约36KDa,10%最底边约25KDa,12%最底部约12KDa。8%胶可 以跑300KDa-36KDa之间的任意蛋白,转膜效率对WB结果的的影响都问题不大。12%,180KDa左右,转膜效率对WB结果的影响都问题不大 (300KDa蛋白如何,没有尝试过)。
电泳一般采用恒流,45mA-55mA(应根据电泳仪器适当调整,要注意仪器最高限压;此条件适合gibco modelV16型,胶宽20cm)。采用恒流的优点,保证最快速度完成电泳(电压会逐渐增大),省时且减少扩散;但是由于电泳速度过快时会发热而溶胶,所以你必须想办法散热。散热不好,条带出现波浪状。
注意事项:
1.聚丙烯酰胺的30%母液会降解,要4度避光保存。
2.APS会失效,10%APS一般保质期才一个月左右。-20度分装长期保存。
3.注意Tris buff的PH值,以及平时所用的水的PH值。PH值改变会使带型非常怪异,所有蛋白和溴酚蓝压成一条细线(即便在分离胶中),如果溴酚蓝前有红色染料,那么此染料彗星式拖尾从溴酚蓝一直延伸到胶底部(溴酚蓝此时涌动极缓慢,位于胶中上部)
4.上下层式电泳装置若漏液,哪边漏液,电泳条带往哪边倾斜。内外式电泳装置漏液,则装置处于短路状态,液体会过热。SDS-PAGE胶可能局部自溶,条带扭曲、变形。
5.配胶用玻璃板和边条应及时洗净。玻板未洗净的坏处很多。尽管玻璃看似平滑,但是一些细微的凹陷处会凝结肉眼无法分辨的胶颗粒(摸上去疙疙瘩瘩),其坏 处是,在该部位极易导致不均匀的胶块,样品经过该处或电泳散热不好,条带变形。
如果是RNA的超薄胶,胶板的颗粒会导致局部巨大气泡,非常难于清除;蛋白 胶也会导致一些小气泡的产生。玻板没洗净的另一个坏处是,由中学物理知识可知,玻璃表面越光滑粘附越牢,未洗净的玻板会削弱和胶体间的粘附力,拔梳子或边 条时会产生微小的错动,胶体下部出现大量气泡(夹在玻板和胶之间,这个关系不大);严重的错动会使上样时,样品从胶和玻璃之间的间隙漏光,或者甚至胶和玻 板分开。为避免拔梳子时的错动,可在电泳缓冲液中拔梳子(比水更好,有SDS润滑)。
判断玻板是否洗干净,用手摸一下有没有疙疙瘩瘩的;最好用洗洁精之类,洗衣粉、肥皂都不好用。
6.上样时,不要把tip深入胶孔过深(或者采用细的尖端拉长的专用上样枪头),可能会错开胶和玻板,样品会泄露。
7.未加样的孔应添加高浓度SDS LB平衡,否则最外侧条带会拉宽变形。通常20ul样品(含5ul undefinedSDS LB),可用8-8.5ulundefinedSDSLB平衡。同理,点marker的lane也要加入同样体积的LB。LB若在室温放置太久和新鲜的在比重上会有差异,在新旧LB靠近的地方,条带会拉宽或挤压变形。因此,同一块胶上,煮样及填空白所用LB应一致。LB应-20度保存。
8.增加上样量不一定会提高荧光信号强度。增加上样量的后果通常只能是让你的内参粘连,所有的蛋白条带都扭曲变形。因为增加上样量最多只能提高几倍,而 WB灵敏度是以10的几次幂量级的,所有目的蛋白信号的唯一方案就是IP富集(可特异提高目的蛋白浓度数百数千倍,并去除其它杂蛋白干扰)。增加上样量的 另一个坏处是,本来高表达的蛋白,诸如内参,在同样WB条件下,可能出现荧光灼烧式粹灭(一晃而灭)或者条带中空。
仍以6孔板80%以上汇合度为例,细胞裂解液和SDSLB通常都是投入200ul(最少80ul,这样面积的培养板如果裂解液投入太少,回收时的损失就太 大,上样就很难做到一致),而这种浓度条件下制备的样品,电泳时上样量要控制在5-6ul,最少2.5ul,最多10ul,10ul时内参基本已经开始粘 连成一条线,带型出现波浪纹;当然并非不可以多上样,再多对WB结果影响不大(没有的仍然没有),且条带都很丑。
9.不同样品上样时,可考虑将样品体积调成一致或类似。如果一组IP样品和一组细胞裂解液样品,前者通常洗脱体积为15ul,刚好+5ul undefinedSDSLB达到常规20ul上样体积;后者第8点提到只上5ul,同样+5ul SDS LB(比重同,不会互相挤压),最好补加10ulDDW使总体积一致。通常这样不同条件获得的样品,中间最好用“空白”泳道隔开(空白仅指无蛋白,仍应加入8-8.5ul undefinedSDSLB),这样才能确保每条带都很美观。
10. SDSPAGE胶配好暂时不用时,需用电泳缓冲液灌满空间(拔去梳子和底边边条后的间隙),用保鲜膜包裹防止液体蒸发,短期室温或稍长期4度保存。个人习 惯为,灌好分离胶用饱和的正丁醇灌满上层,保鲜膜包裹,次日再灌堆积胶。两种方案都可以。所以如果预计次日工作量较大,可以提前灌好SDS PAGE。
样品是否一定需要定量再上样?——不是的,这是浪费时间的一个操作。
我们首先来讨论一下蛋白定量的科学性。
蛋白定量首先需要一个参照系(标准蛋白),参照蛋白通常选择BSA,也就是说你所谓的定量是对应于BSA的浓度的一个参照值。这里面存在一个问题,即忽略 了蛋白折叠和空间构象对光吸收、折射的影响;不同的蛋白折叠和构象还会影响颜料分子的嵌入。因此你其实默认将所有的蛋白等同于BSA在溶液中的折叠状态、 光吸收情况下,然后做出的一个相对判断,这就存在一个“系统误差”——还不算上测量误差等等,就已经很不准确了。
其次,裂解组织或细胞的时候,那些裂解液中,某些溶剂本身会使CoomassieG250或者Bradford法(实际也是Coomassie染色)显 色,尤其是去污剂之类;例如NP40,就会使Coomassie显蓝色,可以完全覆盖掉蛋白与Coomassie作用产生的蓝色。因此,如果你的裂解液里 面含有这类物质,所谓的蛋白定量实际是NP40颜色和蛋白染色的叠加,根本不会符合标准蛋白曲线的线性规律,自然定不准。【需要说明:我目前只知道 NP40会这样,因为我们从来不去定量,没有做过更深入的研究。】
实际上保证你的内参一致,是从样品制备开始的,上节末尾有提到,不赘述。如果你从制备样品时就注意过定量问题,实际上通常内参差别不会太大。如果真的有差 别,通常我们会先跑少量的样品,目的是让Actin或tubulin更清晰、不会粘连、不会过曝光。从而在第二轮跑正式结果前,根据第一轮的内参的荧光信 号做微调,大致能做到相当;最多可能需要第三轮。以上的试验可以精确到0.1ul差异(我很多体外生化试验之前的调整酶量就是这样进行,此时根本不可能有 足够的蛋白让你去定量)。当然凭曝光强弱调整上样量的前提是,你的WB技术很稳定,很可靠,这是需要相当多的经验积累,如果你一时掌握不了,可以让经验更 丰富的师兄师姐或者导师帮忙。
顺便提到,有人问过用Quantity One等定量软件对光密度定量后计算差别从而调整上样体积是否可以?——不可以。因为WB结果经过多重放大,精确计量只代表相对于对照组的相对比值,与实际比值还是有误差,所以按计算值更改上样量仍会出现偏差。
如果非要做定量的话,要注意你的裂解液配方,把每种溶剂都先加入到显色液中尝试一下,避免那些本身会显色的物质出现在你的裂解液中;当然,某些物质的去除可能影响对组织蛋白的提取。所以这是一个两难的问题。
转印——转印效率对WB结果的影响并不如书本和网络上宣扬的那么大!
首先必须澄清的是,很多时候新手会把WB结果无信号归咎于转膜不够充分,实际上转膜效率对最终结果的影响所占比重并不高,真正取决定性因素的是抗体识别能力的强弱,以及如何正确使用抗体,这个问题下节讨论。
有一个简单的例证,Biorad提供的3mm厚滤纸初次使用时,通常转膜效果不理想(从预染的marker深浅可知),但对多数一般丰度的蛋白的检测没有 任何影响(建议少用这种新滤纸做那种含量特别稀少或者转膜非常困难的蛋白)。
没有很好的策略可以解决这个新滤纸的问题;尝试过浸泡(会泡烂的)、预电泳 (会稍微好点)等等,效果均不理想。通常,最初一两次的使用就是用于转一般蛋白。每次用完后清水稍微冲洗一下表面盐分(要控制水流;水流太大,纸张会烂掉 的),晾干再用;只要没冲烂可以一直反复使用。
其次,尽管转膜效率不起决定性作用,但由于WB的流程相对较长,所以每个步骤的叠加作用会被级数放大,因此在试验的每个步骤我们仍会力求更好,因此以下仍会深入探讨如何提高转膜效率。
针对提高转膜的效率,个人认为最先应该考虑到的是仪器的选用。
这里不是歧视“made inChina”,国内的厂家很少注重不断提高自己产品的品质,走低端策略,对于电泳仪这种技术含量不高的仪器倒可以凑合;但说到转膜仪,能把一个简单的 匀场电极做好的还真不多(就我道听途说的,国外厂家为了使电极之间场强更加均匀,那种大块金属板表面是镀过极薄的白金层);因此转膜效率和均匀方面孰优孰 劣不言而喻。
目前,个人使用过的国产电转槽(湿转)唯Tanon仿Biorad的还可以(算替它们免费打个广告吧,Tanon仿bioradmini3型 电泳仪,在某些方面(主要是操作)上还优于Biorad原装;目前我认为“中国制造”的灵魂就是仿造并超越前者);半干转仪一律进口,用过Biorad、 amersham和英国公司的scie-plas。
PS:批评一下Biorad。Biorad的半干转仪,其硬质塑料外壳会莫名其妙的碎裂(绝非摔裂、磕碰,因为底面一般放在桌上后除定期需要清洗去除残留 盐渍,基本不会移动;即便如此它自然就会碎裂),以致于其核心组件未坏的情况下,因外壳碎裂不得不报废该仪器(其外壳里包裹电源线,外壳碎裂,电源则无法 接通,导致仪器报废——我们先后买过3台都是这样的毛病,其间间隔了3年,不能肯定近来Biorad有没有改进这个问题;如果没有,我想市场迟早会被其他 公司所取代)
对这三款产品,Biorad的外壳有明显的质量缺陷,容易过早报废;amersham独特的蜂窝式设计确实对转膜效率有所提高,但蜂窝式使得与“三明治” 的接触面减少,电转时散热不太好,做大蛋白(半干转仪也可以做大蛋白转膜,效率确实不如湿转,但抗体好、蛋白丰度一般时仍可采用)长时程(3hrs)转膜 需要在4度冰柜或cold room进行;英国的产品,价格较高,公司不太出名国内不一定有代理,但质量和散热都非常好。
这里谈到了半干转和湿转。这两种转印方法各有优劣。湿转适合所有的蛋白,转膜效率最佳,但靡费试剂、溶液,对普通分子量大小的蛋白转染操作时间长于半干转 (下文会具体给出条件);半干转适合分子量较小的蛋白,省时、省试剂。蛋白分子大小如何界定呢?
习惯上认为150KDa以上为大蛋白,其他均属于小蛋白, 当然这只是一个相对标准。因此,通常对大蛋白的转印多数人会选择长时程的湿转,那么刚才提到的amersham半干转仪的缺陷实际上没太多实质意义,何况 还可以外加条件弥补;而且用半干转做大蛋白转印本来就是高手在悬崖上跳舞,此时转膜效率是否更高已经没有任何意义——我说过转印效率对WB结果不起决定因 素。
湿转、半干转缓冲液配方请参考分子克隆,有必要指出的是,实际上用半干转缓冲液进行湿转效果也非常理想,通常这种缓冲液可以反复使用多次(<=5次 常规1hr电转,如有3hrs长时程转染,缓冲液使用次数会减少),不过要注意初始电流(同样温度下,初始电流高,表明缓冲液中电解质剩余不多,为确保转 膜温度,可开始或中途更换新鲜转膜液)。
电转液中SDS增加蛋白的水溶性,促进蛋白在电转液中泳动。若不加SDS,蛋白会沉淀在胶上,但SDS过多会影响 蛋白与膜的吸附。甲醇能使SDS与蛋白分离,甲醇浓度越高SDS与蛋白分离越快,但高浓度的甲醇对蛋白会有固定作用,不利于蛋白从胶里跑出来。一般来说甲 醇的浓度为20%,但PVDF膜截留marker上交联的小分子颜料的能力很差,可考虑提升甲醇的浓度至25%(它对普通蛋白的转膜影响不太,颜料截留更 多);大蛋白转印可考虑降低甲醇浓度,另外SDS必须添加。
甲醇的另一个作用是降温,旧的电转液由于电解质的消耗和甲醇的挥发,电转时电流或电压变化更 快、温度升高也更快,因此可考虑增加额外的甲醇;这点对半干转缓冲液的意义较大,因为半干转缓冲液消耗很慢,缓冲液放置的时间较久,甲醇挥发比较严重,可 临时少量补加。
湿转一般采用稳压,电压控制在120-140V,时长1-3hrs(小蛋白控制在1hr,大蛋白3hrs),有人会选择60V过夜,从实验的效率来讲太浪 费时间,对转膜到底有多大程度的增益难说,不太推荐;半干转一般稳流,依milliporePVDF膜附带说明书,当2.5-3mA/cm2,电压 25V(biorad半干仪额定值不能超过25V;amersham半干转仪,限压30V。
通常跑不到这么大的电压,除非是新的滤纸或很久的转膜液,或常 温放置的转膜液,或超大的胶),时间一般15–45min(常规用0.5hr);如果是3mA/cm2,时间不要超过0.5hr。这两款仪器的限压要 求,可能出于保护仪器的目的;amersham为防止过载,当电压>30V,~35V附近的时候,保险会自动跳掉、切断电转仪电源。至于UK的,即 使整张大板胶室温电转3hr以后,最大电压仍为18V(因此足见其散热性能的优良)。注明:以上半干转仪时间的要求是针对PVDF膜的,这里面有一个很有 趣的问题。
尽管厂家一再表明PVDF膜比NC膜对蛋白的截留更高(但NC带负电性,理论上它的吸附量应该更高,所以通常同样使用TBST这种低盐洗液背景 比PVDF膜高),但PVDF膜对小分子的截留明显不如NC膜,因此交联在预染marker上的颜料小分子很容易穿过PVDF膜,造成转膜过度的假象(除 可考虑提升甲醇浓度至25%外,也可以考虑增加marker的上样量);NC膜没有这种问题,所以通常它的转膜时间也是小蛋白1hr、大蛋白3hrs,电 流控制在200-300mA(16cundefined9cm大胶300mA,如果胶面积小很多可以考虑降低,实际上相当于2.5-3mA/cm2左右)。
NC膜唯一不 如PVDF膜的地方,在我看来是它的强度:干燥的膜易碎。当然目前某些厂家为解决这个问题,将NC成分涂在另一种介质的表面,这种膜的支持强度和PVDF 膜类似,但转膜效率稍差于纯NC膜,且有明显的正反面;因此制作转印三明治的时候要特别注意正反面。此外,20KDa以下的蛋白宜采用0.22uM孔径的 转印膜,但也不是一定必须。
对于大蛋白转印,很多书本建议采用低浓度胶,如6%SDS-PAGE。但实际操作发现,6%太软,制作转印三明治的操作非常麻烦;且内参如Actin、 tubulin都在45KDa附近,而6%胶底边溴酚蓝指示60KDa左右,除非采用坛子上有人提过的灌不同浓度的胶或者梯度胶,否则需要同时跑两块胶, 因此也不是很推荐。个人经验认为300KDa以下8%胶(底边36KDa)都可以胜任,可以适当调整选择7-8%胶可以兼顾内参和大蛋白。
转膜最好在低温进行(高温会局部溶胶或降低转膜效率),尽管很多半干转仪没有具体要求,但用预冷过的电转液湿滤纸比未预冷的转印效率更高。因此,有条件建 议湿转、半干转均在低温(4度)进行。此外,湿转缓冲液可以考虑转印前-20度预冷,时间不能长于2hrs,否则电泳液冻结;mini3型有内置冰槽的, 冰槽置-80度预冷。
制作转膜三明治的过程中,书中强调过滤纸、胶、膜的大小最好一致,否则没有胶、膜隔开部分的滤纸会因为直接接触而产生局部短路,降低内阻增加电(压)流, 造成升温过快。但进口仪器随厂原包装附带的滤纸有限,如果每次都切割新滤纸,消耗过快、初次使用的膜转膜效率不高,且增加额外的操作。
因此,实际上滤纸大 一些也不太要紧,但是我会选择已经切割好的和胶面积最接近的滤纸;通常膜的大小会严格控制在与胶面积一致或略小一点点。操作时在保证每一层均无气泡的前提 下,叠好后再碾压几个来回,力道要均匀、恰好;力量过大,胶会被拉伸,条带会扭曲、变形。
碾压的目的不是为了驱赶气泡(完全可以做到叠加的时候每一层都无 任何气泡;诸如采用多层薄滤纸时可以一张张的叠加),更重要的作用是让膜和胶贴得更紧密。可以理解的是,对于蛋白分子的大小而言,胶和膜之间的间距是数十 万倍计的,因此更紧密的接触无疑是转膜成败的关键。
胶和膜需不需要泡呢?不少公司实验讲座时提出泡胶、泡膜,实际操作中没有发现泡和不泡的转膜效率有多大差别。实际上,胶只需要在电转液中浸一下即可(可以 减少与滤纸或者膜之间的气泡);而膜也只要湿掉沾上电转液即可,同样多沾些缓冲液会有利于减少气泡(PVDF要先用甲醇湿润再投入缓冲液;NC直接进缓冲 液)。
如何监控转膜的效率呢?
在早期,鄙人亦依据分子克隆的介绍,采用立春红染膜。只是后来发现此操作不仅增加额外的操作时间,而且不能说明任何问题,于是抛弃之。首先,立春红(也包 括考染胶)的灵敏度和HRP-ECL体系的灵敏度相差太远,即使立春红染膜无颜色,也无法提示WB结果会不会失败(考染胶无颜色也不能说明转膜充分了,除 非你银染),你仍然得继续后面的操作;相反靶蛋白区域有颜色,亦无法提示靶蛋白就转膜完全了,也许只是另外一个分子量大小相近的蛋白。
因此,无论立春红染 膜失败或成功,你都必须进行后续的操作。而立春红染胶不仅增加额外的操作时间,而且增加一轮漂洗的操作,对膜和膜上的蛋白都不好。
那么为什么不采用更直观 的预染marker做转膜监控的依据呢?理论上,同时转膜、颜色是交联到蛋白上的,因此颜色有多深表明有多少蛋白转印到膜上,非常直观、不会增加任何额外 的操作(固定marker的上样量非常必要)。
当然上面提到针对PVDF膜,由于对小分子截留的局限,颜料分子会在高强度的转印过程中从交联的蛋白上脱落 并穿过膜(颜料与蛋白交联得过深会使整个lane糊掉;太紧(多)可改变蛋白构象使迁移率改变。所以多数情况下颜料和蛋白之间是一种不稳定的交联,这意味 着在某些条件下,颜料会从交联的蛋白上脱落,又因为膜分子孔径以及膜对不同物质吸附能力的差异,颜料小分子会轻易穿过膜到滤纸背面,而蛋白则被牢固的截留 在膜上),造成转膜过度的假象。
现在你知道有这种局限,要么更换NC膜;要么采用上面提到的非常稳定的转膜条件、注意必要的细节,就可从根本上忽略掉其对 转膜效率的指示,而把预染的marker仅仅作为一个分子量大小的指针,当然同时可确认之前的转膜操作无误(没有插反电极,放反膜),也可为后续抗体孵育 操作时膜的正反面做必要的提示。
不同公司预染的marker效果不太相同,一般NEB和invitrogen的常规分子量预染marker要上8-10ul,MBI的只要5ul(胶大小20×30cm),Biorad-mini3型(8.2×7.4cm)MBI最低2.5ul转膜后就足够清晰。
抗体孵育——WB真正的难点:抗体的使用是一种艺术,一种抗体一个脾气。
对WB结果有决定性影响的因素是抗体的效价和如果正确使用手中的抗体。无论你如何提高自己的转膜技术,高效和低效之间往往只有数倍的差异,而抗体的效价可以差数千倍以上;同样,正确使用抗体可以保证抗体效价不被过分的拮抗,谋求特异性和非特异性背景之间差异的最大化。
封闭最短可以缩减到5min:
很多人把高背景或者黑板,认定为封闭不充分,其实这也是没有吃透WB(说实话,经常看到坛子上很多人这样给人答疑,非常的无语);实际上封闭(极端点)也 是个可有可无的步骤,真正对降低背景起核心作用的是抗体稀释液中的组分。当你的抗体使用次数较多时,基本不用封闭也可以有较低的背景;如果抗体很新,其实 封闭最短可以缩减到5min(没试过更短的,不一定不可以)。
那什么导致高背景甚至黑板呢?抗体的质量不太好(主因),或者抗体稀释液的配方有问题(增大 抗体稀释比略微有所改善)。封闭液的配方可以和抗体稀释液相同,也可以不同;通常封闭液是最简单(单方)的抗体稀释液,而抗体稀释液可能是复方。
封闭很少出状况。不过在milk做封闭剂的封闭液中,milk颗粒未完全溶解时,微小的颗粒会粘附在膜上增加星点状背景。
紧接封闭操作的是抗体孵育,之间不要用TBST漂洗。抗体孵育成败的关键首要在抗体本身的质量,包括抗体的效价和背景的强弱,然而历来商品化的抗体质量参 差不齐。各家生产的抗体中质量问题最严重的是scbt的抗体,但它们的价格却是最便宜的。
其实,scbt在美国的售后服务还是相当不错的,只要按他们的要 求操作仍然能举证抗体的质量问题,可以更换、交换(你指定的他们销售的另外一种抗体),有时候还会附送一些ProAbeads之类的,所以不奇怪它的普及 率;也正因为此,如果你参考文献的话很容易找到这家公司的。
可惜,在国内代理商不可能提供相应的服务,因此购买他们的商品存在不小的风险。所以,通常我更 推荐sigma和Abcom、R&D等。诸如Sigma公司销售的Actin,cat.A5316,cloneAC-74,Mab更是作为新手练 习专用的抗体;它的背景很低,效价很高,可以1:30,000稀释(是普通一抗效价的30倍)。
此外,尽管cellsignaling是做磷酸化抗体起家 的公司,但它们的抗体的质量总体感觉也不好,所以有其它公司可以选择时,我们会刻意避免购买cellsignaling的产品,诸如Akt总抗和 Ser473(这两种抗体R&D的产品效价不错)。
(特别注明:前几年CST的AKT抗体识别的条带在55KDa左右,而其他公司均认定为60KDa;最近还有战友提问这个抗体,重新检索了一下,应该是原 AKT抗体(包括磷酸化)已经被CST自己下架了,从目前的几个抗体示意图看,大小也已与其他公司一致;因此,要不要买取决你自己)
抗体公司在出售抗体的时候,在广告上会玩很多花样。A.不给整张膜标记的图示。很多抗体质量不好,背景高杂带多,如果靶蛋白区域较干净时,他们通常只给很 窄的一条靶蛋白区域的图例。B.不给内源性蛋白标记的图示,用IP的样品取代。
通过IP可以富集抗原,减少杂蛋白,通常很容易拿到背景很干净,信号很强的 图例。C.用制备抗体时高表达的多肽做阳性样品,不给全长蛋白的图例。由于多肽可以IP富集,且不存在空间折叠的问题(通常裸露在外),因此很容易标记。 除以上花样外,不少抗体用途上还有局限,因此挑选抗体时要睁大眼睛。
此外,有一点值得注意的是,scbt销售的抗体表面上很多(通常1ml,而别的公司通常是200ul或者100ul),然而实际上它的浓度也仅为 0.2λ,所以实际上他们抗体的质量也和其他公司的没区别都是200ug(常规)。所以在早期,我一直强调如果固定即用型一抗的浓度(稀释的一抗)为 1ug/ml时,scbt的抗体最佳稀释比为1:200。
不过近来发现,实际上他们的抗体1:2K稀释效价也不错,因此也许其他公司的一抗可以进一步提高 稀释比。现在判断当时scbt抗体之所以采用1:200稀释,实际上是因为自制的ECL不够灵敏,所以建议用灵敏性更好的ECL,这样可以减少抗体的消耗 量,进一步降低实验成本。
一抗通常4度过夜,效率较常温1-1.5hrs高;二抗通常1:5K稀释,室温0.5-1hr(过久并不会明显提高信号强度甚至会减弱(相当于洗膜),同 时造成高背景进一步影响特异与非特异荧光信号强度间的对比。
孵育时间过久还会迫使你延长TBST的时间,这对抗原识别能力较弱的抗体更致命)。洗膜通常用 TBST,也可以考虑高盐洗膜液(NaCl0.5M);洗膜时间一般10miundefined3或者5miundefined4。抗体孵育和洗膜时,摇床速度不能过快也不宜过慢,需 要简单摸索一下。
聊完以上那些浅显的基本常识,接下来谈我认为最麻烦的:
首先,抗原抗体识别原理的物理学解释,抗体孵育过程实际上就是一个竞争结合的动力学过程。
由于抗原和抗体特异性结合的效率是非特异性结合的数百万——数十 亿倍,所以当特异性抗体与非特异性‘抗体’(不好描述,指添加在抗体稀释液中的封闭蛋白,我们可以把它们看成非特异性的一抗)竞争时,尽管抗体稀释液中封 闭蛋白的浓度是一抗浓度20K:1以上,在碰撞几率相同的条件下,由于时间的积累加孵育过程中反复漂洗(孵育抗体要摇晃,对吧),特异性一抗积累了识别、 结合优势。
同样在TBST等漂洗的情况下,由于亲和力等原因,使得一抗很牢靠并进一步增加对比优势,再经过2抗特异性积累和ECL的最终放大形成特异性条 带和非特异性背景之间荧光信号的巨大差异。
由以上碰撞结合、亲和力差异这些基本现象,我们可以解释WB中出现的现象。
1.在样品制备章节中,我强调PBS漂洗去培养基中BSA的作用。当时只描述了不漂洗的后果,是在BSA位置任何抗体都可以非特异性的结合从而显影。用碰 撞的原理去解释:当杂带很浓的时候,抗体的特异性已经毫无意义,它只符合物理定律中的碰撞几率原理;因为你的杂带浓,碰撞几率大,粘附一抗的机会多,因此 它就会显出来,但切记这是杂带。当杂蛋白含量高到一定程度,“任何”一种一抗都可以在该位置显出条带。
因此,用条带的深浅去判断是否是靶蛋白,这是非常不科学的。在默认抗体有效的情况下,如果抗原靠近区域没有杂带,背景干净,依据分子量大小可以初步确认靶 蛋白的位置;如果有杂带,并不一定是特异性条带就比杂带亮,也许杂带蛋白浓度大,非特异性粘附的一抗更多。目前判断杂带和靶蛋白的唯一标准是分子量;而当 分子量大小类似时,只有通过增加过表达或IP纯化的样品做阳性对照,或者KD该蛋白的样品做阴性对照一起电泳转膜,才能准确区分靶蛋白和杂质。(用IP或 KD样品也是验证商品化抗体是否有效的可靠方案)
2.WB结果白板(无信号)和黑板(高背景)。从抗体孵育角度解释白板和黑板问题(当然也可能与ECL显影有关,此点下一节有阐述),白板主要是可能是抗 体本身效价不高,或者抗体稀释液中封闭蛋白过多,竞争结合时封闭蛋白完全挤掉了特异性的一抗,特异性条带和非特异性背景同样结合强度——无信号。
黑板,也 主要是因为抗体效价不高,而抗体稀释液中封闭蛋白的拮抗作用又没有完全发挥,此时无法有效识别抗原的一抗会等效的粘附转印膜的任何位置,显影后整张膜全 黑。
有时候膜接近全黑,但可以若有如无的观察到靶蛋白,出现问题的原因虽然仍是抗体效价不高,封闭蛋白不能完全起到作用,不过此时可以通过改变抗体稀释液的配方,提高特异性和非特异性结合的信号差异,降低背景。
“这点非常重要”——当你的抗体标不到抗原时(白板),先弄出条带(黑板),再解决黑板(高背景)问题。第一步应该是通过倍比降低一抗的稀释比(从 1:1K降到1:500到1:250到。。。;注:二抗稀释比不要轻易改变,增加二抗浓度对WB结果不会有太明显的改变),摸索到一个可以标出信号的条 件。
此时,由于一抗浓度过高,背景可能非常高,但背景的问题可以再调整,有没有特异性条带却没办法改变;实在没有靶蛋白的信号,只能更换别的公司生产的抗 体。
那么如何通过改变抗体稀释液的配方,寻求抗体孵育的最佳条件呢?
首先要认清,抗体稀释液没有一个绝对通用的配方;一种抗体一个脾气。
其次,抗体稀释液的配方要兼顾与特异性抗体竞争抗原的强度,以及对其他区域的封闭强度两方面的平衡。
目前,抗体稀释液中可以充当封闭蛋白的主要有milk,BSA和gelatin,似乎很少的样子。。。可是,你有没有考虑过“复方”?——这下配方无限多了吧。
1.采用milk,BSA和gelatin的单方。3% milk,5%BSA,<=0.4%的gelatin,并根据实际情况改变(若出现白板,请降低封闭剂浓度。;黑板多加)。gelatin大家可能比较陌生,不过如果你翻翻抗体公司卖给你的原装抗体里面液体的配方,你会发现很多抗体溶液里都有它。
2.很多情况下单方的浓度很高了,背景问题仍不能解决。那么请尝试两两或者三种不同比例混合。不过有些细节还是要自己摸索的,比如gelatin+BSA时,gelatin浓度不能无限提高(会完全竞争掉抗原抗体特异性结合,导致白板),BSA的浓度较随意。
3.抗体稀释液可用低盐浓度的TBST,也可用高盐浓度的缓冲液(NaCl 0.5M)以降低背景;但是切记,有的抗体对盐浓度敏感,包括构象和溶解度,因此需要尝试。
如果以上策略仍然无法解决高背景,可将稀释好的一抗先跟平时实验剪切下来的membrane的边角料(新的)放在一起孵育个把小时再用。如果还不行,彻底无解,结论抗体太差;请更换别的公司或者抗别的区段的同种抗体。
这三种封闭蛋白的差异:
milk和gelatin封闭效果不错且价格便宜,但milk容易发霉变质,且国产milk脱脂不充分亦增加背景、拮抗抗原抗体结合(拮抗与抗原结合尤其 突出;更不知道三聚氰胺会不会影响抗原抗体结合或者ECL!?);BSA封闭效果不佳,且价格较昂贵。个人推荐gelatin,gelatin主要成分是 骨胶原,蛋白很大,但可以通过加热打断成小蛋白,从而实现封闭各种分子量大小的蛋白。
通常,我会将gelatin加到TBST中后直接灭菌,灭菌过程中, 不溶的gelatin会逐渐溶解;由于gelatin可以灭菌(milk不可以),所以同样条件下,gelatin的存放时间明显长于milk。
实际上, 抗体可以反复用很久(通常一般的抗体常规稀释比,可以连续用一个月以上;对于老手用过2、3次的旧抗体更prefer,因此背景降到很低而效价正是最高的 时候),通常抗体最后失效的原因不是因为使用次数过多,而是染菌;比较旧的抗体都会有很多的沉淀在底部,更长时间还会有异味。稀释的一抗靠添加NaN3并 于4度保存延长使用时间;二抗不能加NaN3,但放在-20度反复冻融延长使用寿命(抗体并非绝对不可以冻融;只是高浓度的原始管抗体的冻融对效价会有较 大影响)。
抗体稀释液(一抗、二抗稀释液也可同也可不同)和封闭液可以相同,可以不同,因为抗体稀释液可以采用复方的,所以即使混合了少许,不会对抗体的使用产生较大的影响。不过封闭液用milk时,gelatin稀释的一抗混入少许milk会影响其使用寿命。
由于以上诸多可变因素,实在无法总结出一种万金油式的抗体稀释液(只能是多数通用),因此对待具体问题时,请具体对待(多花点心思摸索吧)。
原装抗体的保存:
1. 分装。每次一管,但是可能太多占地方,而且还会有粘壁的损失,不是很推荐。
2.1:1加甘油,冻于-20度,可长期保存,此方法实际上也被很多国内的试剂公司广泛采用,诸如博士得,中杉,它们小包装的进口抗体通常都是1:1添加 甘油保存于-20(不信去查查)。值得注意的是:不是所有的抗体都可以1:1添加甘油,要以抗体说明书为准,如果说明书中本身存在甘油,或特殊注明外,通 常可以采用。
显影——淬灭的祸因
当抗体孵育、TBST漂洗结束后,进入ECL显影步骤。当然,目前国内一些实验室已经换用仪器扫描荧光信号,而我们更是二抗直接连荧光蛋白连ECL和常规的底片显影都省略了,因此以下所讨论的某些细节问题可能不再出现。
首先坛子上还有少数实验室处于DAB显色时代,奉劝一句,都走到最后一步了就不要节省了;DAB显色的灵敏性是远远不够的。目前较为流行的仍是化学发光法,下面简述一下其原理:
交联在二抗上的HRP酶能够特异水解过氧化物产生化学能;ECL中主要包含两种成分,催化剂和中间递质(白色瓶子主要是过氧化物如双氧水)。催化剂的作用 不用废话了,其中间递质主要是吸收化学能并将其传递到luminol之类的化合物上产生电子跃迁从而激发荧光。因此催化剂的效率和中间递质的传递效率直接 影响荧光的强弱。本人曾自制过ECL并尝试改良,其间发现很多化合物、金属离子如Fe、Cu等能催化并高效传递能量,且不依赖于HRP浓度,荧光信号能瞬 间曝强;这其实与下文提到的一些粹灭原因有联系。
当初我自制的ECL最大的毛病在于稳定性不好,同理,商品化的ECL同样也有保质期的问题,往往最先失效的组分就是过氧化物,尽管他们必然添加过抗还原 剂。其实,检验ECL是否失效的方法很简单,取稀释的即用型二抗1-2ul加至ECL混和液中观察荧光强度,即可知道ECL是否失效。
商品化的ECL价格并不便宜,因此ECL孵育的最佳方案是:将ECL滴加在保鲜膜上或干净的支持物上,如废弃的旧底片、塑料盒,有机玻璃盒等等(注意,国 产的保鲜膜很多质量不过关,有两种潜在的问题,下面详述),然后倒扣膜于保鲜膜上,夹住一个角来回拖动数次至ECL均匀(有的产品说明书要求孵育 1min)。另外平铺一张干净的保鲜膜,将膜揭下控干ECL、倒扣包好,然后显影。mini3型胶整张膜孵育,ECL总体积0.7-1ml足矣。
哪些因素会影响荧光信号的强弱呢?(这部分实际上与导致荧光淬灭的因素部分重合)
1.保鲜膜的质量。a.国产的保鲜膜,很多厂家偷工减料打价格战,造成保鲜膜很薄亦破损。ECL滴加到保鲜膜上容易从肉眼不易分辩的小孔泄露,一方面 ECL反应不充分,另一方面ECL所含液体会溶解试验台上残留未知的化学药品颗粒(也许你或其他人曾经不小心打翻过某种化合物)、灰尘等等,造成荧光淬 灭。在简述原理时,我曾经提到过很多化合物、金属离子能直接催化过氧化物水解,在极短的时间内消耗掉所有的HRP酶,荧光转瞬即逝。
b. 保鲜膜的化工原料或拉制过程中,残留未知的化学试剂,引起荧光淬灭;廉价的保鲜膜问题尤多。
目前,国产的就“妙洁”比较过关,保鲜膜厚度和化学物残留都不影响ECL显影。
如果买不到合格的保鲜膜,也可以用其他物品替代;但要特别注意这些支持物表面的干净,最好不要有任何化学试剂残留,包括风干的TBST盐结晶颗粒。
2.ECL孵育结束后膜需要控干。中学物理学过水会吸收光谱,因此任何液体包括ECL溶液本身都会干扰荧光强度,所以ECL孵育结束时需要控干。一般夹起 膜,让膜的一角或一边接触吸水纸;但是请注意不能让膜完全干掉。某些信号较弱的ECL,膜彻底干掉后荧光会消失;较好的试剂盒,荧光相对稳定,但完全吸干 以后,背景荧光也会升高,而且会严重影响重新标记新抗体时的背景。因此,按照我的方法,让自由流下的多余的ECL被吸走就好。
3.控干的膜要仔细用保鲜膜包被,防止接触外界异物造成荧光淬灭;同时,包裹保鲜膜时要细心,尽量不让正面保鲜膜和膜之间出现皱褶。皱褶的地方会堆积多余的ECL,要么形成一条荧光的亮线;要么由于液体吸收荧光或者局部区域HRP过度消耗,呈线条状淬灭。
4.在膜放入压片夹之前,最好肉眼观察一下荧光信号的强弱,这有利于判断实验可能出现的问题。很多人提问看见很强的荧光了,但怎么压片都没有信号,那么就 是快速淬灭(闪灭,再怎么快速操作也拿不到这种情况下的荧光信号)造成的;有这个观察至少能够判断ECL孵育之前实验操作应该问题不大,而问题主要是淬 灭。如果你省略这个步骤,那问题就非常复杂了,因为之前任何操作都可能导致白板,根本无法判断。
除上述的问题以外,还有哪些因素会导致荧光淬灭呢?
1.抗原浓度太高,荧光信号过强,快速淬灭。在电泳的章节就曾提过,如果上样量太大,不仅条带会粘连、弯曲,更可能导致淬灭。因为局部区域过多的HRP酶 会快速消耗底物呈富氧态,条带出现烧灼样(取出膜会发现一条明显的暗黄色的带),荧光信号会闪灭或者条带空心(条带灰黑不实,则可能是显影液接近失效)。
2. 抗原浓度太低,荧光信号太弱,相对慢一点的淬灭。可能第一次压片能够获得很弱的条带,随后都不可见。
3. ECL试剂质量。除过氧化物因接触空气被逐渐还原外,ECL的成分通常都不稳定。如呈递电子跃迁能的中间递质其化学性质就不太稳定,因为它转换能量的特性决定其必是一个有中间态的化合物,长期接触空气会被逐渐氧化失效;luminol也有保质期。
4. 操作时间过长,膜逐渐彻底干掉。商品化的ECL会出现波形渐变,开始信号逐渐增强,背景一起升高;随后荧光完全衰减淬灭。
5.不良的实验习惯。坛子上曾经有人问及淬灭问题,提到显影夹子很脏。脏的显影夹主要有两种污染,铁锈和不慎沾染的显影液、定影液,前者造成闪灭(催化作 用),后者可能直接导致无荧光;只要有任何失误,诸如不小心刮破保鲜膜,保鲜膜包裹不严实(其实通常还真的不严实,因为你不可能包裹好几层)。于是,我问 他用一个很脏的东西不觉得很别扭嘛,为什么不先抄起抹布擦干净再用呢。
6.淬灭的假象。少量沾在膜表面的二抗也会激发荧光,但ECL孵育时稍微晃动就消失了,可能产生淬灭的错觉。
Stirpping:
完整的WB流程结束后,可能需要重新标记某些膜,这时候就“可能”需要对膜stripping。这里用了“可能”二字,即不一定必须,那么首先讨论一下stripping的必要性。
Stripping排除其他的不讲,整个流程会耗费不少的时间,从节约时间的角度来讲能不做最好;并且stripping条件过于激烈或多轮洗脱时,还会 剥离已转印到膜表面的抗原使WB的最终信号削弱;此外,strippingbuffer未清除干净时,其中的某些组分一定会干扰第二种抗体的结合。那么在 什么条件下不需要stripping呢?
1. 当A蛋白和B蛋白分子量差别5KDa以上(10KDa左右更保险),可将膜分割开(分别标记不同的抗体),这样就不需要 stripping膜重新标记,且一次就能得到想要的结果。有人会将两种抗体混在一起标记整张膜,这样操作的前提是非常熟悉这两种抗体的杂带位置,且各自 的杂带都不会干扰目的蛋白才可以如此操作;并且这样的抗体下次使用有了局限性,因此不太推荐。
2. 两种抗体种属来源不同时,如鼠抗、兔抗等等,即使蛋白位置非常靠近无法切膜,这时候也不一定需要stripping。轮番标 记这两种不同的抗体时,只需要用TBST涮掉表面的ECL,时间可长(一时腾不出手来就扔在那边一直漂,中间可以换换液)可短(换液涮两三次),然后直接 用第二种抗体标记同一张膜。不过,如果其中一种蛋白显影太强,隔天显第二种蛋白时可能会出现干扰(可能强可能弱);但只要两个蛋白不是同样大小或者连在一 起了,可以在最后作图时用PS切出来。
3. 当两种抗体种属来源相同时,有一种情况也不需stripping,诸如标记某蛋白的磷酸化和总蛋白时。由于磷酸化抗体通常识别能力较弱(相对于总蛋白而言),信号一般都不太强,此时也仅需简单的漂洗;但标记的顺序一定是先标磷酸化后标总蛋白。
以上避免stripping主要从节约时间的因素考虑,实际上当上述2、3点存在时,个人还是会用stripping buffer简单漂一下,然后用TBST换液漂洗3次再上第二种抗体;但如果时间较紧,就会真的省略。
Stripping膜至少有三种方法:
A.用TBST一直洗膜,洗8hrs以上就可以清除多数蛋白的信号,信号过强的蛋白如内参除外。所以,当你的抗体非常弱的时候,孵育过夜实际上相当于stripping,新的信号不会出现,原来的信号也会被漂洗干净,出现白板。
B.请参考millipore PVDF膜附送的说明书配方(没有,请Google),上面提到如果信号很强,可以在50度反应。需要提醒的是,由于巯基乙醇亦挥发,可以考虑临时添加。
C.如果做过IP,你就知道可以用低pH的Glycine-HCl洗脱beads上特异性吸附的蛋白。同 理,0.1MGlycine(pH2.5,30min)也可以用于stripping膜,这是已知最廉价的高效洗膜方案。而且经过 TBST(10miundefined3)漂洗后,膜上的pH早就恢复到正常值,此时不存在上述strippingbuffer残留成分干扰第二种抗体标记的情况。如果 第一种蛋白信号过强,同样可以在50度stripping提高洗膜强度。
此外,膜风干后,表面结合的二抗也会脱落,不过不是很推荐。
看完stripping这个部分,再多想一层,你一定会找到同一张膜多次标记不同抗体的窍门。
如果有A、B、C三种蛋白,因分子量过于接近无法切膜:
a.如果A、B的抗体同是兔抗,C的抗体为鼠抗,则最合理的标记顺序为A-C-B或者B-C-A。
b.A、B、C的抗体同种来源,A、B几乎在一起,条带粘连;C靠下。如果A、B重要性差不多、B信号稍弱,则B-C-A;如果A的结果是最关键的,尽管 B信号弱,一般还是A-C-B。因为,经过两次strippingbuffer洗脱,且两轮WB流程的时长都超过8hrs(一抗习惯4度过夜孵育),实际 上在标记第三种抗体时,相对于经历了3次不同强度的stripping;不要担心第一种抗体还会有很强的干扰。
如何正确使用Quantiy One定量:(以下文字系摘录+改写整合)
WB做完,有时候我们需要对结果进行定量。目前最备受推崇的应该非Biorad公司的QuantiyOne软件莫属,然而其$6K左右的售价令很多人望而 却步;不过,这款软件之所以敢如此漫天要价,其最大的优点莫过于自动识别条带代替人工分析以降低主观误差,同时它的很多功能渗透了艰深的数理理论以及概率 统计的原理,这在其它各类免费的随机附送的蛋白定量软件上是乏善可陈的。不过,令人扼腕的是,又有多少人真的只是把它当做一款普通的“免费”软件在使用着 ——暴殄天物。
目前多数免费软件主要采用的是等高线定量法(VolumnContour),QuantiyOne也包含这部分内容且在网络教程里面流传最广,以至于很多 人只会这种并不准确的蛋白定量方法。等高线定量法通过半自动描绘电泳条带的等高线边缘来得到等高线区域内部面积,再将该面积乘以区域内平均光密度值得到条 带内部总的信号量。
这种分析方法有明显的弊病:人为圈定背景值且假定不同泳道的背景亮度一致;等高线的绘制处于“半自动”状态,即需要人为判断作为等高线 标准的电泳条带的边缘;最致命的是在几个电泳条带距离十分接近的时候几乎无法绘制单一条带的轮廓(常出现连续的几个条带等高线相连而无法分离出单独条带的 轮廓)。
实际上,最科学、严谨的蛋白定量方法应该是泳道/条带轨迹定量法(TraceTracking)。这种方法使用起来步骤较为繁琐,必须通过泳道识别— 电泳条带识别这两个连续的步骤才能进行定量。但它最大优点在于可以完全抛弃人为主观因素进行全自动定量。
他的定量方式为:首先根据不同电泳条带的光密度值 绘制光密度曲线,然后计算光密度曲线下面积作为电泳条带的定量根据。其次,它能够最大程度的排除不同泳道之间的背景差异,让各个泳道上的不同电泳条带在一 条几乎相同的起跑线上进行对比;这个背景排除功能是等高线法无法做到的。最终,采用泳道/条带轨迹定量法分析条带光密度后,再结合Gauss Model Bands对电泳条带进行数理统计。
执行步骤:
由于Quantiy One只能识别黑白的TIF文件,而实际上发表论文时,很多杂志也要求提供TIF等格式的图片而非JEPG格式的;因此,考虑到这两层因素,我建议大家从一开始扫描就统一存贮为TIF格式的图片。
1.创建泳道:
a.打开我们要分析的电泳图片(以蛋白质电泳为例)。
b.选择“Lane-Auto Frame lanes”。这时如果您的电泳图片比较标准的话,Quantity One就会自动识别出每条电泳泳道的位置,而且将各个泳道的路径用一条红线标画出来;
如果您对软件自动标记的泳道不甚满意,您可以通过“Lane-Edit Frame”下级各个子菜单提供的功能对泳道框架位置、大小等进行调节;调节方法就是通过鼠标的拖放来实现。
如果您只需要分析其中几个泳道的数据而不想其他泳道的标记干扰您的视线,您可以通过“Lane-Single Lane-Remove Lane”删除您不需要的泳道的标记。
注意:
不是所有的电泳图片的泳道都能够被Quantity One自动识别。在不能自动识别的情况下,QuantityOne就会弹出对话框告诉您它不能识别泳道。这时大家就需要手工绘制电泳泳道。方法和上面一样,同过“Lane-Single Lane-CreatLane”来实现。只要用鼠标在您的电泳图片上顺着待标记的泳道的中轴线拖放就可以绘制出该泳道的标记红线。
2.不同泳道的背景排除:
我们要将我们的电泳图片进行前面谈到的泳道识别,不管是自动方式还是手工识别。然后选择“Lane-LaneBackgroud…”命令。选择该命令 后,鼠标即变成一个绿色的“+”,将鼠标移到您打算进行背景排除的泳道,点击左键一下,立刻就会弹出如下图所示的对话框。
其中“OpticalDensity”显示得是该泳道光密度值的分布情况。大家会注意到这个分布曲线并不是紧贴着纵轴,而是位于纵轴上方分布。造成这个 “悬空现象”的原因就是该泳道自身存在着一定的光密度“背景”。这个背景的存在导致该泳道上各个电泳条带之间不能处于同一水平进行对比;如果考虑到相邻的 其他泳道更是因为不同背景的存在而无法对比不同泳道上的不同电泳条带。
如何去除这个背景呢?我们继续看右边的“Lane BackgroudSubstruction”对话框。这个对话框的上方“All Lanes”表示可以对所有泳道进行一次性处理;下方的“SelectedLane”表示仅针对此次选中的这个泳道进行处理。我们用鼠标选择 “Selected Lane”的“LaneOn”选项。然后在下面的“Rolling Disk Size”里填上“5”,再按“回车”键。大家再来看看现在“OpticalDensity”中出现了一条紧紧沿着光密度曲线分布的酱色曲线。这条曲线将 泳道的背景与电泳条带的光密度分布十分精确的划分开来。
注意:
“Rolling Disk Size”是什么意思呢?实际上就是数理里面的微分等差逼近。我们可以将QuantityOne提供的去除背景功能想象成是一个滚动的小球。如果这个小球 的半径越小,那么它沿着光密度曲线滚动时滚过的路径就越发精细,具体反映在酱色曲线的轨迹越发靠近黑色光密度分布曲线的基线。
因此从理论上来说 “Rolling DiskSize”的取值越小,它的去除背景效果越好。大家可以试着选择一个较大的值比如80,再来看看此时酱色曲线的轨迹。当然也不能取太小的值。因为 如果取值太小,大家可以想象这个十分微小的球就会滚入光密度曲线内部,造成有效阳性信号的损失。个人感觉5~20是一个比较好的取值范围。
我们可以对每条泳道依葫芦画瓢进行以上类似的背景排除工作。不过有时候如果大家觉得可以使用同样的标准,即相同的“Rolling DiskSize”进行排除,那么我们可以在“Lane Backgroud Substruction”对话框中选择上方的“All LanesOn(same level)”选项。然后在“Rolling DiskSize”中填上一个合适的值,点击“Done”按钮确认就可以了。此时该电泳图片上所有的泳道都以相同的标准进行了背景去除。
3.创建条带:
前面我们已经识别和分析了电泳图片的泳道,下面我们开始研究电泳条带的问题。首先在这里明确两个名词翻译:“Lane”指电泳泳道;“Band”指电泳条带(在下文中一律使用这两个英文名词来表示这两个意思)。
a.电泳条带的识别:和前面研究Lane一样,首先我们要对Lane上的Band进行识别。识别方式同样分为自动识别和人工识别。自动识别的时候选择“Band-Detect Bands…”命令。这时Quantity One会自动弹出一个“Detect Bands”的对话框。
注意:
第一次用QuantityOne时,您的Bands识别以后仅仅是画出了一条红色的粗线,而在上面的演示图片中却是上下括号的形式。其实大家可以在 “Band-BandAttributes”选项中设置Bands的显示形式。在下方的“Style”选项卡中选择“Brackets”就会将原来的粗线 改为括号形式了。
b.如果要自动识别所有Lanes上的Bands就钩选Lands后面的“All”选项;如果仅仅想自动识别某一条Lane上的Bands就钩选“One”,然后在后面的输入框中填上您想识别的泳道号码;
如果仅仅想识别泳道上部分光密度值最强的Bands,可以在“Bands”后面的选项中选择“Limit”,然后填上相应的数目(比如10)。Quantity One就会仅识别该泳道上光密度最大的10条Bands;
如果您发现自动识别的Bands宽度太窄,圈定Band的红色括号内范围小于黑色的光密度影。此时可以使用“LaneWidth”命令来调整Bands识 别的宽度。用鼠标点击“LaneWidth”后面的向上箭头就会发现红色括号逐渐变宽,最后要求其范围略为宽过其包绕的Band影迹;
如果您发现自动识别功能没有识别您想要的Band或者误识别了您不想要的Band,您还可以使用上面的工具栏进行“添加”或“删除”。将您的鼠标指向每个图标,Quantity One就会知道弹出一个黄色的提示信息告诉您这个按钮的功能。
4.高斯建模与结果分析:
a.高斯建模,即用数学方法(微积分)对正态分布曲线进行统计分析。理论上每道band的光密度都呈正态分布的平滑曲线(每个band中间位置蛋白压缩最 紧浓度最高,向上向下因扩散而减弱);然而实际操作过程中,如果荧光过强,往往出现类似方波的曲线(曲线无peak峰,上方为平滑直线,超识别的最高阈 值);荧光较弱宜出现波浪样(多)波峰,或难与背景区分。
首先电脑会统计band中每个点的光密度值,并对这些值按正态分布进行归类,当读值不符合正态分布的原理时,电脑会自动做出选择以对偏离正态分布的数据进行统计学取舍(置信区间),从而最终优化光密度曲线呈正态分布。
如何进行高斯建模呢?很简单,只要执行“Band-Gauss Model Bands…”命令就可以了,执行后QuantityOne回弹出一个对话框问您要对那条泳道进行高斯建模。请钩选“One”,然后再输入需要建模的泳道代码就可以了;如果选择了“All”则对所有泳道进行高斯建模,当然建模这么多泳道会费一点时间了。
b.观察结果:执行完高斯建模以后使用“Band-Band Information”命令来观察结果。
方法是将变成蓝色惊叹号的鼠标移到刚才已经识别的Band的部位,一个详细的“BandInformation”信息框就会立刻弹出来。在这幅图中我们必须注意的是“Trace”和/或“Gauss ModelTrace”,因为我们刚才辛苦了半天就是为了得到这个数据。这个数据表示的就是Band光密度分布曲线下面积,也就是QuantityOne用来表达Band内分子总量的方式。
最后,我们计算、记录的值,应该是“Gauss Model Trace”这一栏的读值。