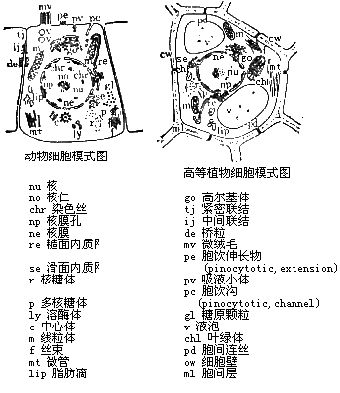
细胞质
互联网
细胞质(cytoplasm)又称胞浆是由细胞质基质、内膜系统、细胞骨架和包涵物组成。
(一) 细胞质基质
细胞质基质又称胞质溶胶(cytosol)是细胞质中均质而半透明的胶体部分,充填于其它有形结构之间。细胞质基质的化学组成可按其分子量大小分为三类,即小分子、中等分子和大分子。
小分子包括水、无机离子;属于中等分子的有脂类、糖类、氨基酸、核苷酸及其衍生物等;大分子则包括多糖、蛋白质、脂蛋白和RNA等。细胞质基质的主要功能是:为各种细胞器维持其正常结构提供所需要的离子环境,为各类细胞器完成其功能活动供给所需的一切底物,同时也是进行某些生化活动的场所。
(二)内膜系统
内膜系统(endomembrane system)是通过细胞膜的内陷而演变成的复杂系统。它构成各种细胞器(organelle),如内质网、线粒体、高尔基复合体、溶酶体等。这些细胞器均是互相分隔的封闭性区室,各具备一套独特的酶系,执行着专一的生理功能。
1、内质网 (endoplasmic reticulum,ER)是扁平囊状或管泡状膜性结构,它们以分支互相吻合成为网络,其表面有附着核糖核蛋白s体者称为 粗面内质网 (rough endoplasmic reticulum,RER),膜表面不附着核糖核蛋白体者称为 滑面内质网 (smooth endoplasmic reticulum,SER),两者有通连。
核糖核蛋白体附着在内质网上,其主要功能是合成分泌蛋白质( 如免疫球蛋白、消化酶等),但也制造某些结构蛋白质(如膜镶嵌蛋白质、溶酶体醇等)。粗面内质网分布于绝大部分细胞中,而在分泌蛋白旺盛的细胞(如浆细胞、腺细胞),粗面内质网特别发达,其扁囊密集呈板层状,并占据细胞质很大一部分空间。一般说来,可根据粗面内质网的发达程度来判断细胞的功能状态和分化程度。
滑面内质网多是管泡状,仅在某些组胞中很丰富,并因含有不同的酸类而功能各异,
①类固醇激素的合成,在分泌类固醇激素的细胞中;滑面内质网膜上有合成胆固醇所需的酶系,在此合成的胆固醇再转变为类固醇激素;
②脂类代谢,小肠吸收细胞摄入脂肪酸、甘油及甘油一酯,在滑面内质网上酯化为甘油三酯,肝细胞摄取的脂肪酸也是在滑面内质网上被氧化还原酶分解,或者再度酯化;
③解毒作用,肝细胞的滑面内质网含有参与解毒作用的各种酶系,某些外来药物、有毒代谢产物及激素等在此经过氧化、还原,水解或结合等处理,成为无毒物质排出体外;
④离于贮存与调节,横纹肌细胞中的滑面内质网又称肌浆网,其膜上有钙泵,可将细胞质基质中的Ca2+泵入、贮存起来,导致肌细胞松弛,在特定因素作用下,贮存的Ca 2+ 释出,引起肌细胞收缩。胃底腺壁细胞的滑面内质网有氯泵,当分泌盐酸时将CI ˉ 释放,参与盐酸的形成。
2、高尔基复合体(Golgi complex)由扁平囊、小泡和大泡三部分组成,它在细胞中仿分布和数量依细胞的类型不同而异。扁平囊(saccule) 有3-10 层,平行紧密排列构成高尔基复合体的主体,它有一面常凸超称生成面(forming face),另一面凹陷,称成熟面(maturing face)扁平羹上有孔穿通,并朝向生成面。生成面附近有一些小泡(vesicle),直径为40~80nm,是由附近粗面内质网芽生而来,将租面内质网中合成的蛋白质轻运到扁平囊,故小泡又称运输小泡。
大泡(vacuole)位于成熟面,是高尔基复合体的生成产物,,包括溶酶体、分泌泡等。
溶酶体逐渐离开高尔基复合体而分散到细胞各部。分泌泡互相融合,其内容物电子密度增高,成为分泌颗粒。在蛋白质分泌旺盛的细胞中高尔基复合体发达。高尔基复合体对来自粗面内质网的蛋白质进行加工、修饰、糖化与浓缩,使之变为成熟的蛋白质,如在胰岛B细胞中将前胰岛素加工成为胰岛素。
高尔基复合体具有多种糖基转移酶,许多蛋白质在此被糖化形成糖蛋白。此外,名种溶酶也在高尔基复合体浓聚形成初级溶酶体。
3、 溶酶体(lysosome)为有膜包裹的小体,内含多种酸性水解酶,如酸性磷酸酶、组织蛋白酶、胶原蛋白酶、核糖核酸酶、葡萄糖苷酸和脂酶等,能分解各种内源性或外源性物质。它们的最适ph为5.0。
不向细胞中的溶酶体不尽相同,(但均含酸性磷酸酶,故该酶为溶酶体的标志酶。
按溶酶体是否含有被消化物质 (底物)可将其分为初级溶酶体(primary lysosme)和次级溶酶体(secondary lysosome)。
(1) 初级溶酶体: 也称原溶酶体(protolysosome) 。 一般呈圆形或椭圆形,直径多介于25~50nm近年发现亦有长杆状或缓状溶酶体。其内容物呈均质状,电子密度中等或较高不含底物。在少数细胞,如破骨细胞和炎症部位的中性粒细胞,溶酶体酶可被释放到细胞外发挥水解作用。
(2) 次级溶酶体:也称吞噬性溶酶体(phagolysosome),是由刺级溶酶体和将被水解的各种吞噬底物融合而构成,因此其体积较大,形态多样,内容物为非均质状。 根据其作用废物的来源不同,分为自噬性溶酶体和异噬性溶酶体。自噬性溶酶体(autophago lysosome)的作用底物是内源性的,即来自细胞内的衰老和崩解的细胞器或局部细胞质等。
异噬性溶酶体(heterophago lysosome)的作用底物是经由细胞的吞饮或吞噬而被摄入细胞内的外源性物质,是溶酶体与吞噬体融合而成,多见于吞噬了细菌的中性粒细胞和吞噬了异物的巨噬细胞。并噬性溶酶体与自噬性溶酶体中的底物有的被分解为单糖、氨基酸等小分子物质。
它们可通过溶酶体膜进入细胞质基质,被细胞利用;有的则不能被消化(如尘埃、金属颗粒等异物、衰老细胞器的某些类脂成分),它们残留于溶酶体中,当溶酶体酶活性耗竭,溶酶体内完全由残留物占据,则称之为残余体(residual body)。
在哺乳动物,残余体滞留在细胞中,常见的残余体有脂褐素颗粒和髓样结构。均由自噬性溶酶体演化而来。脂褐素颗粒(lipofuscin granule)为不规则形,由电子密度不同的物质及脂滴构成,在光镜下呈褐色,多见于神经细胞、心肌细胞、肝细胞及分泌类固醇激素的细胞,并随年龄增长而增多。
髓样结构(myelin figure)的内部为大量板层排列的膜,可能因膜性成分消化不全所致。初级溶酶体与吞饮小泡或其它小泡融合形成多泡体(multivesicular body),其外有界膜,内含很多低电子密度小泡,基质具有酸性鳞酸酶活性。
3、中间丝(intermediate filament)又称中等纤维,直径约为8~11nm,介于细丝与粗丝之间,因而得名。中间丝可分为五种,各由不同蛋白质构成。在成体中绝大部分细胞仅含有一种中间丝,故具有组织特异性,且较稳定。五种中间丝的形态相仿,难于分辨。但用免疫组织化学方法则能将它们区分,从而可进一步分析细胞的类型。
(1) 角质蛋白丝(keratin filament):分布于上皮细胞,在复层扁平上皮细胞内尤其丰富,常聚集成束,又称 张力丝 (tonofilament)。张力丝附着于桥粒(一种细胞连接),能加固细胞间的连接。张力丝除起支持作用外,还有助于保持细胞的韧性和弹性。
(2) 结蛋白丝(desmin filament):分布于肌细胞,在横纹肌细胞内,结蛋白丝所形成的细网连接相邻肌原纤维并使肌节位置对齐;在Z膜股处,细网包围肌原纤维并与细胞膜连接。在平滑肌细胞内,结蛋白丝连接在密体与密斑之间形成立体网架,并与肌动蛋白丝相连。总之,结蛋白丝作为肌细胞的细胞骨架网,发挥固定和机械性整合作用。
(3) 波形蛋白丝(vimentin filament):主要存在于成纤维细胞和来自胚胎间充质的细胞。在少数含有两种中间丝的细胞中,波形蛋白丝是其中的一种,波形蛋白丝主要在核周形成网架, 对核起机械性支持,并稳定其在细胞内的位置。
(4) 神经丝(neurofilament):存在于神经细胞的胞体与突起中,由神经丝蛋白组成,与微管共同构成细胞骨架,并协助物质运输。
(5) 神经胶质丝(neurogial filament):主要存在于星形胶质细胞内,由胶质原纤维酸性蛋白组成,多聚集成束,交织走行于胞体,并伸入突起内。
4、微梁网(microtrabecular lattict)是用超高压电镜等技术在完整细胞中观察到的由直径3~6nm的纤维交织形成的立体网架。有人认为它是一种镶嵌在其 它纤维系统中的微梁网格。也有人认为,它是微管、微丝和中间丝系统紧密联系和交错相插,或是某些被磨损的细胞骨架所显示的图像。总之,它仍是一个有争议的结构。
4、线粒体(mitochondria) 常为杆或椭圆形,横径为0.5~1ηm,长2~6 η m但在不同类型激胞中线粒体的形状、大小和数量差异甚大。电镜下,线粒体具有双层膜,外膜光滑,厚6~7nm,膜中有2 ~ 3nm小孔,分子量为1万以内的物质可自由通过;内膜厚5~6nm,通透性较小。外膜与内膜之间有约8nm。膜间腔,或称外腔。由膜向内折叠形成线粒体嵴(mitochohdrial crista),嵴之间为嵴间腔,或称内腔,充满线粒体基质。基质中常可见散在的,直径25~50nm。电子致密的嗜饿酸基质颗粒(matrix granule),主要由磷脂蛋白组成,并含有钙、镁、磷等元素。基质中除基质颗粒外还含有脂类、蛋白质、环状DNA分子核糖体。线粒体嵴膜上有许多有柄小球体,即基粒(elementary particle),其直径为8~10nm,它由头、柄和基片三部分组成。球形的头与柄相连而突出于内膜表面,基片镶嵌于膜脂中。
基粒中含有ATP合成酶,能利用呼吸链产生的能量合成ATP,并把能量贮存于ATP中。细胞生命活动所需能量的约95%由线粒体以ATP的方式提供,因此,线粒体是细胞能量代谢中心,线粒体嵴实为扩大了内膜面积,故代谢率高,耗能多的细胞。嵴多而密集大部分细胞的线粒体嵴为板层状。杆状线粒体的嵴多与其长轴垂直排列,圆形线粒体的嵴多以周围向中央放射状排列;在少数细胞,主要基分泌类固醇激素的细胞(如肾上腺皮质细胞等),线粒体峭多呈管状或泡状;有些细胞(如肝细胞)的线粒体兼有板层状和管状两种。
线粒体另一个功能特点是可以合成一些蛋白质。目前推测,在线粒体中合成的蛋白质约占线粒体全部蛋白的10%,这些蛋白疏水性强,和内膜结合在一起。线起体合成蛋白质均是按照细胞核基因组的编码辑导合成。如果没有细胞核遗传系统,线粒体RNA则不能表达。因此表明线粒体会成蛋白质的半自主性。
关于线粒体形成的机制,较普遍接受的看法是,线粒体依靠分裂而进行增殖。线粒体的发生过程可分为两个阶段,在第一阶段中,线粒体的膜进行生长和复制,然后分裂增殖。第二阶段包括线粒体本身的分化过程,建成能够行使氧化磷酸化功能的机构。线粒体生长和分化阶段分别接受两个独立遗传系统的控制,因此,它不是一个完全自我复制的实体。
5、过氧化物酶体(peroxisome)又称微体(microbody),是有膜包裹的圆形小体,直径为0.2 ~ 0.4μm,多见于肝细胞与肾小管上疫细胞。在人其内容物为低电子密度的均质状;在某些动物尚含电子致密的核心,是尿酸氢化酶的结晶。过氧化物体含有40多种酶,不同细胞所含酶的种类不同,但过氧化氢酶则存在所有细胞的过氧化物酶体中。各种氧酶能使相应的底物氧化,在氧化底物过程中,氧化酶使氧还原成过氧化氢,而过氧化氢酶能使过氧化氢还原成水。这种氧化反应在肝、肾细胞中是非常重要的。
6、核糖体(ribosme) 是由核糖体RNA(rRNA)和蛋白质组成的椭圆形致密颗粒,并非膜性结构,(因属细胞器,故在此叙述)颗粒大小约为15nm×25nm。核糖体由一个大亚基与一个小亚基构成。大亚基合两条rRNA与约40个相关蛋白质分子,并有一条中央曾;小亚基含一条rRNA与约40个相关蛋白质分子,非功能状态的核糖体单个存在。当一定数量(3~30)的核糖体由一条mRNA细丝穿行于它们的大、小亚基之间把它们串联起来,则成为功能状态的多核糖体(polyribosome), 电镜下呈串珠状或花簇状。核糖体能将mRNA所含的核苷酸密码翻译为氨基酸序列,即肽链合成的肽链从大亚基中央管释出,肽链可进一步聚合形成白质细胞质基质中的游离核糖体(free ribosome)合成细胞自身的结构蛋白,如细胞骨架蛋白细胞基质中的酶类等,供细胞代谢、增殖和生长需要。因此,在旺盛增殖中的细胞游离核糖体极多。于内质网膜表面的附着核糖体(attached ribosome)除合成结构蛋白外,主要合成分泌性蛋白。核糖体丰富的细胞,光镜下胞质呈嗜碱性。
(三)细胞骨架
细胞的特定形状以及运动等,均有赖于细胞质内蛋白质丝织成的网状结构―细胞骨架(cytoskeleton)。细胞骨架是由微管、微丝、中间丝和微梁网组成。
1、微管(microtubule)是细而长的中空圆柱状结构。管径约15nm,长短不等,常数根平行排列。微管由微管蛋白(thbulin)聚合而成。微管蛋白单体为直径约5nm的球形蛋白质,它们串连成原纤维,13条原纤维纵向平行排列围成微管。微管有单微管、二联微管和三联做管三种类型。细胞中绝大部分微管为单微管,在低温、Ca2+ 和秋水仙素作均下易解聚为微管蛋白,故属于不稳定微管。二联微管主要位于纤毛与精子鞭毛中,三联微管参与构成中心体和基 体,均为稳定微管。
微管具有多种功能。微管的支架作用可保持细胞形状,如血小板周边部的环行微管使其呈双凸圆盘状,神经细胞的微管支撑其突起,如果加入秋水仙素使微管解聚,则血小板变圆,神经细胞突起缩回。微管参与细胞的运动,如细胞分裂时,由微管组成的纺锤体可使染色体向两极移动,如果加入秋水仙素则分裂停止于中期,纤毛和鞭毛的摆动、胞吞和胞吐作用、细胞内物质的运送都需要微管参与。
2、微丝(microfilament)广泛存在于多种细胞中,微丝常成群或成束存在,在一些高度特化的细胞(如肌细胞),它们能形成稳定的结构,但更常见的是形成不稳定的束或复杂的网。它们可根据细胞周期和运动状态的需要,改变其在细胞内的形态和空间位置,并能够根据在细胞的不同状态而聚合或解聚。
分布于肌细胞和非肌细胞中的微丝分细丝和粗丝两种。细丝(thin filament)直径约6nm,长约lμm,主要由肌动蛋白(actin)组成,故又称肌动蛋白丝(actinfilament),通常所说的微丝指此而言。细胞松弛素B能使细丝解聚,从而抑制细胞运动;粗丝(thick filament)直径侧10~15nm,长约1.5μm,主要由肌球蛋白(myosin)组成,故又称肌球蛋白丝(myosinfilament)。
微丝是肌细胞内的恒定结构。在横纹肌细胞内;细丝与粗丝以一定比例(约为2:1)有规则排列成肌原纤维,其收缩机制已明确。平滑肌细胞内细丝与粗丝之比约为15:1,二者的排列不规则。非肌细胞中一般只能看到细丝,粗丝可能因存在时间短暂,或于电镜标本制备过程中解聚为肌球蛋白,难于观察到。在某些因素作用下,非肌细胞中的微丝迅速解策为其结构蛋白;在相反因素作用下,结构蛋白又装配成微丝。其中细丝交联成网以构成细胞骨架的一部分,并维持细胞质基质的胶质状态;细丝与粗丝的局部相互作用能引发运动。在活跃运动的细胞(主要在细胞质周边部)或细胞局部(如伪足),以及需察机械支持的部位(如微绒毛),都有丰富的微丝。因此,微丝除具有支持作用外,还参与细胞的收缩、变形运动、细胞质流动、细胞质分裂以及胞吞、胞吐过程。
3、中间丝(intermediate filament)又称中等纤维,直径约为8~11nm,介于细丝与粗丝之间,因而得名。中间丝可分为五种,各由不同蛋白质构成。在成体中绝大部分细胞仅含有一种中间丝,故具有组织特异性,且较稳定。五种中间丝的形态相仿,难于分辨。但用免疫组织化学方法则能将它们区分,从而可进一步分析细胞的类型。
(1) 角质蛋白丝(keratin filament):分布于上皮细胞,在复层扁平上皮细胞内尤其丰富,常聚集成束,又称 张力丝 (tonofilament)。张力丝附着于桥粒(一种细胞连接),能加固细胞间的连接。张力丝除起支持作用外,还有助于保持细胞的韧性和弹性。
(2) 结蛋白丝(desmin filament):分布于肌细胞,在横纹肌细胞内,结蛋白丝所形成的细网连接相邻肌原纤维并使肌节位置对齐;在Z膜股处,细网包围肌原纤维并与细胞膜连接。在平滑肌细胞内,结蛋白丝连接在密体与密斑之间形成立体网架,并与肌动蛋白丝相连。总之,结蛋白丝作为肌细胞的细胞骨架网,发挥固定和机械性整合作用。
(3) 波形蛋白丝(vimentin filament):主要存在于成纤维细胞和来自胚胎间充质的细胞。在少数含有两种中间丝的细胞中,波形蛋白丝是其中的一种,波形蛋白丝主要在核周形成网架, 对核起机械性支持,并稳定其在细胞内的位置。
(4) 神经丝(neurofilament):存在于神经细胞的胞体与突起中,由神经丝蛋白组成,与微管共同构成细胞骨架,并协助物质运输。
(5) 神经胶质丝(neurogial filament):主要存在于星形胶质细胞内,由胶质原纤维酸性蛋白组成,多聚集成束,交织走行于胞体,并伸入突起内。
4、微梁网(microtrabecular lattict)是用超高压电镜等技术在完整细胞中观察到的由直径3~6nm的纤维交织形成的立体网架。有人认为它是一种镶嵌在其 它纤维系统中的微梁网格。也有人认为,它是微管、微丝和中间丝系统紧密联系和交错相插,或是某些被磨损的细胞骨架所显示的图像。总之,它仍是一个有争议的结构。
(四) 中心体
中心体(centrosome)多位于细胞核周围,由一对互相垂直的中心粒(centriole)构成。中心粒呈是短圆筒状,长0.5μm直径为外0.2μm,由9组三联微管与少量电子致密的均质状物构成其壁。相邻的三联微管相互斜向排列,状如风车旋翼。在壁外侧有时可见9个球形的中心粒卫星(centriolar satellite)。大小约70nm。在细胞分裂时,以中心粒卫星为起点形成纺锤体,参与染色体的分离(详见“细胞周期”)。有纤毛或鞭毛的细胞,中心粒形成基体,参与微管组的形成。
(五) 包涵物
是细胞质中本身没有代谢活性,却有特定形态的结构。有的是贮存的能源物质,如糖源颗粒、脂滴;有的是细胞产物,如分泌颗粒、黑素颗粒;残余体也可视为包涵物。
1、糖原颗粒(glycogen granule)是细胞贮存葡萄糖的存在形式,于PAS反应时呈红色。电镜下,其电子密度高,无膜包裹,并呈两种类型:β颗粒,直径为20~30nm,形状不规则,分散存在。多见于肌细胞;α颗粒,是β颗粒的聚合体,呈花簇状,大小不一,多见于肝细胞。
2、脂滴(fat drop)是细胞贮存脂类的存在形式,内含甘油三酯、脂肪酸、胆固醇等。脂滴在脂肪细胞中最多,其次为分泌类固醇激素的细胞。在前者,常常一个脂滴即占据细胞的绝大部分空间;在后者, 则多是小的球状。在普通光镜标本制备过程中, 脂滴被二甲苯、乙醇溶解而遗留大小不等的空泡。电镜下,脂滴无膜包裹,多是低或中等电子密度,与所含脂肪酸的不饱和程度有关。
3、分泌颗粒(secretory granule)常见于各种腺细胞、内含酶、激素等生物活性物质。分泌颗粒的形态、大小及在细胞内的分布位置因细胞种类而异, 但都有膜包裹。