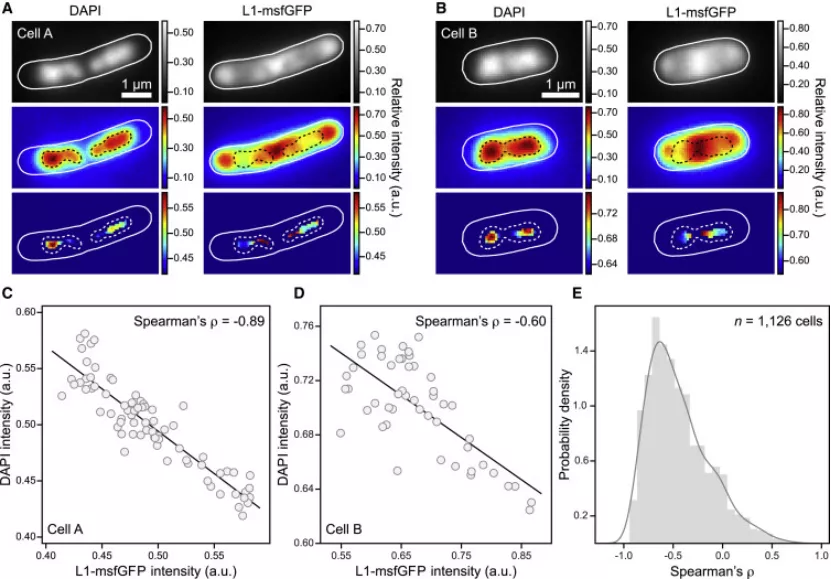
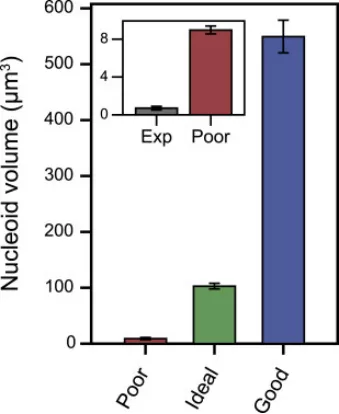
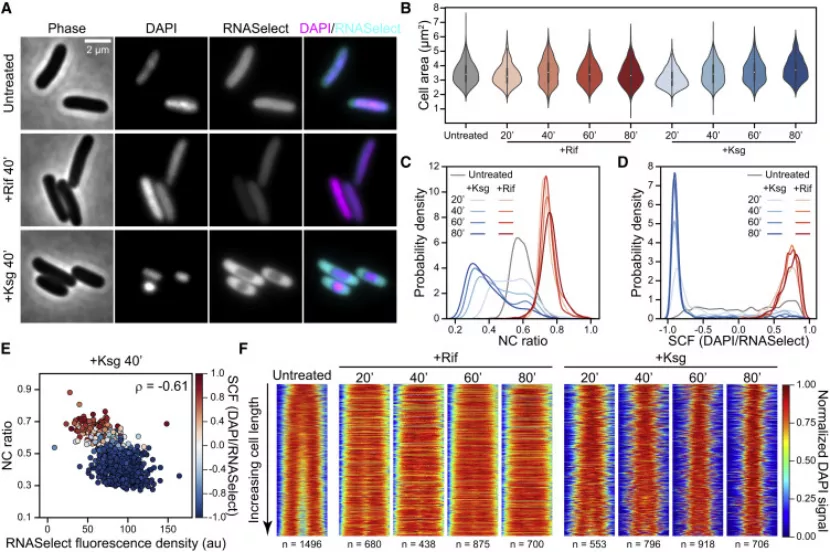
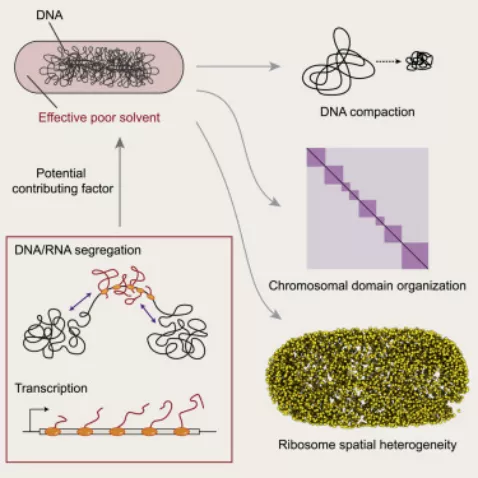
丁香学术2021-09-09
相关产品推荐
Single Cell WGA Kit(MDA),阿拉丁
¥6897.90
WP2uvrA大肠杆菌 (Ames试验)毒理实验 AMES细菌回复突变试验 CA染色体畸变试验 基因突变试验
¥2500
促销活动| 修饰/标记引物主题季,四重惊喜重磅来袭,首单最低可享0元购!
¥0.80
RNApure Tissue&Cell Kit,阿拉丁
¥1197.90
C6 Whole Cell Lysate,阿拉丁
¥2269.90
相关问答
问
请问膜片钳的whole cell模式中的神经元细胞是否包含了轴突(axon)部分
在包涵体蛋白溶解后的重折叠实验中,有什么方法可用于重折叠稀释?
如何确定蛋白是否正确折叠
相关方法
透析得可溶的重折叠σ32 单体实验
2024-05-14
模式动物的麻醉操作与处死方法
2023-03-16
基因插入位点和模式
推荐阅读
Cell 重磅 | 刘光慧等揭示靶向古病毒复活或可实现衰老干预
染色体模式图 idiogram
模式生物大肠杆菌简介