




聚而不散:是什么推动了新物种的进化?

以植物为食的昆虫在两个方面有别于地球上的其他生物。首先,它们非常多样化:每四种昆虫中就有一种是食草昆虫。其次,大多数昆虫专精于特定类型的植物,而排除了它们栖息地的所有其他植物。
对草食性昆虫显著的多样性和专业化的一种解释是,对新的寄主植物的适应常常驱动了新物种的形成。为了验证这一假设,我们必须在有助于宿主适应的性状和阻止物种间成功繁殖的性状之间建立直接联系。
为了研究宿主适应性和物种形成之间的关系,我们关注了一对近亲松锯蝇(北美松树的常见害虫),它们攻击的松树种类不同。松林新二prion pinetum(白松锯蝇)只攻击白松,而红头松锯蝇新二prion lecontei(红头松锯蝇)攻击其他松林品种,但一般不会攻击白松。

我们知道,这两个物种可以——而且偶尔也会——交配。我们在同一时间同一地点发现了它们,我们甚至在野外收集了杂交幼虫。我们还在实验室中进行了杂交,产生了可繁殖的杂交后代。然而,尽管偶尔进行杂交,这些物种在野生环境中在基因、行为和形态上仍然是截然不同的。
总之,我们的观察告诉我们,有些东西在阻止菠萝和lecontei合并成一个单一的物种。在实验室中,这些物种之间的早期杂交暗示了一种可能的解释。我们注意到,当杂交雌性将卵产在松树上而不是白松上时,它们孵化得很好。然而,当杂交雌性在白松上产卵时,针叶干燥,卵无法孵化。换句话说,一些关于杂交雌性的产卵行为是“关闭的”。

为了产卵,雌新双prion用锯齿状结构(产卵器)在寄主针上刻出带有单个卵囊。在此过程中,他们面临两个挑战:绕过保护松针的树脂(这种树脂会淹死松针)和避免针叶干燥(这种树脂也会杀死松针)。值得注意的是,白松的针叶相当薄,树脂含量也比母松柏所偏爱的针叶少得多。因此,我们预测,松果体雌性应该具有能最大程度减少干燥风险的特征(例如,针刺伤更少或更小),而lecontei雌性应该具有能规避宿主树脂的特征(例如,更深的切口可以有效地排出树脂)。
与我们的预测一致的是,我们发现松果树的雌性比lecontei的雌性每根针下的蛋更少。松果体的雌性也从不在针的底部割一个树脂吸水的“前光”(一个没有卵的切口),而N. lecontei雌性总是这样做。我们还发现,松果林雌性比lecontei雌性拥有更小、更直的“锯子”,这表明这些雌性能做更浅的针割。这些性状与我们的预测相符,松果体性状应将干燥风险最小化,而仙人掌性状则有望减少树脂流向鸡蛋(但增加干燥风险)。
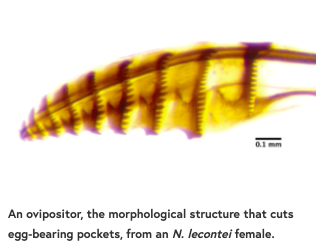
但是这些杂种的特征是什么呢?我们发现,杂交雌性对白松木有强烈的偏好(比如松果木雌性),但与松果木雌性相比,它们有更大的锯子,每根针下更多的蛋,切割更多的压针。这种不幸的性状组合导致了针叶干燥和孵化失败,大大降低了杂交雌性成功繁殖的能力。杂交雌性的下蛋能力差,这可以部分解释为什么尽管在野外偶有杂交,lecontei和N. pinetum仍然是不同的物种。

我们的工作证明了亲本利用寄主特征的不适应组合是如何导致杂交个体的适应度降低的,这反过来又限制了亲本物种之间的基因流动。这为宿主适应促成新物种的形成提供了直接证据。此外,我们的研究表明,在不同寄主上产卵的适应性可能是物种形成的一个特别重要(但目前尚未得到充分重视)驱动因素。








