




弧菌(Vibrio)是菌体短小,弯曲成弧形,尾部带一鞭毛的革兰氏阴性菌。如霍乱弧菌。弧菌属(Vibrio)广泛分布于河口、海湾、近岸海域的海水和海洋动物体内。目前弧菌有91种医学教育|网搜集整理。主要鱼贝类致菌为:溶藻弧菌、鳗弧菌、副溶血弧菌、创伤弧菌、鳗弧菌等;有些弧菌也能引起人类疾病如:溶藻弧菌、创伤弧菌等。弧菌
蓝细菌(Cyanobacteria)旧名蓝藻或蓝绿藻,是一类进化历史悠久、革兰氏染色阴性、无鞭毛、含叶绿素和藻蓝素(但不形成叶绿体)、能进行产氧性光合作用的大型原核微生物。蓝细菌分布极广,普遍生长在淡水、海水和土壤中,并且在极端环境(如温泉、盐湖、贫瘠的土壤、岩石表面或风化壳中以及植物树干等)中也能生长,故有“先锋生物”的美称。蓝细菌有固氮作用蓝细菌与水体环境质量关系密切医学教育|网搜集整理,蓝细菌在污水处理、水体自净中 ...
肺炎衣原体 肺炎衣原体的原体形态多样,电镜下可呈现典型的梨形,其长轴长0.44μm,短轴长0.31μm,平均直径为0.38μm.核区呈圆形,位于细胞中央,平均直径为0.24μm,核区和细胞膜之间有较宽的原生质区医学教育`网搜集整理。肺炎衣原体不能体外培养,只能在细胞内寄生,鸡胚对其不敏感,因此一般不用鸡胚传代,而用细胞培养传代,肺炎衣原体敏感的细胞株为HEP-2或H-292,离心能促进肺炎衣原体对细胞的感染。肺炎衣原体包涵 ...
酪酸梭菌酪酸梭菌又称:酪酸杆菌,酪酸菌。拉丁文:Clostridium butyricum.是人的正常肠道菌之一,医|学教育网搜集整理严格厌氧,为革兰阳性菌。由于产生的酪酸又叫丁酸,所以也被称为丁酸梭菌,丁酸杆菌或丁酸菌。本菌存在于土壤、动物和人体的肠道中。
加德纳杆菌为革兰氏阴性细小杆菌,常呈球杆状,有时呈丝状和多形状,常见两极染色。仅0.4~0.6毫微米至1~2毫微米大小,无活动力、无鞭毛、无芽孢。许多菌株有荚膜医学教育|网搜集整理。在人工培养时必须给予新鲜血液才能生长繁殖,故有“嗜血”之称。但其生物特性与嗜血杆菌不尽相同。加德纳杆菌
人类免疫缺陷病毒(HIV),顾名思义它会造成人类免疫系统的缺陷。1981年,人类免疫缺陷病毒在美国首次发现。它是一种感染人类免疫系统细胞的慢病毒(Lentivirus),属反转录病毒的一种。至今无有效疗法的致命性传染病医学教育|网搜集整理。该病毒破坏人体的免疫能力,导致免疫系统的失去抵抗力,而导致各种疾病及癌症得以在人体内生存,发展到最后,导致艾滋病(获得性免疫缺陷综合症)。艾滋病病毒
原始红细胞简介:正常骨髓原红细胞比例较少,仅占0~2%.在红血病、红白血病中增多,医。学教育网搜集整理并常伴有形态异常;在骨髓增生异常综合征(MDS)、巨幼细胞贫血、急性造血功能停滞、溶血性贫血等疾病也可增多。原始红细胞
白细胞旧称白血球。血液中的一类细胞。白细胞也通常被称为免疫细胞。人体和动物血液及组织中的无色细胞。有细胞核,能作变形运动。白细胞一般有活跃的移动能力医学教育|网搜集整理,它们可以从血管内迁移到血管外,或从血管外组织迁移到血管内。因此,白细胞除存在于血液和淋巴中外,也广泛存在于血管、淋巴管以外的组织中。白细胞
单核细胞(monocytes)是体积最大的白细胞。其细胞核常偏位,呈多形性,如卵圆形、肾形、马蹄形、不规则形等,常有折叠感;染色质呈疏松网状,着色较浅。胞质较多,嗜碱性医学教育|网搜集整理,但因含大量细小的嗜天青颗粒而染成灰蓝色,颗粒含过氧化物酶。血涂片,Giemsa染色。单核细胞
明串珠菌指的是一类不产生孢子,G+C含量低于50%的革兰氏阳性细菌,可以在厌氧或有氧条件下生长,通常表现为过氧化氢酶阴性,不水解精氨酸,其细胞大小为0.5~0.7x0.7~1.2cm.在自然生长环境中,明串珠菌细胞呈对或短链状排列;在某些竞争性生长的环境中,细胞排列成较长的链医学教|育网搜集整理。明串珠菌大多数菌株的最适生长温度在20~30℃之间,异型乳酸发酵,可以进行柠檬酸代谢。明串珠菌
精子分动物精子与植物精子。动物有性生殖过程中的雄性细胞,雄性动物的生殖细胞,异配生殖中的雄配子,由精子器产生的单倍体生殖细胞。而生活中精子更多是指男性成熟的生殖细胞医学教育|网搜集整理,在精巢中形成。精子形态
在微生物界,同样存在类似动植物界的食物链一样的关系。“捕食”细菌的生物,正是科学家们研究微生物的一种强有力的工具:噬菌体(Phage)。噬菌体是感染细菌、真菌、放线菌或螺旋体等微生物的细菌病毒的总称,作为病毒的一种,噬菌体具有病毒特有的一些特性:个体微小;不具有完整细胞结构;只含有单一核酸。噬菌体基因组含有许多个基因医学教育|网搜集整理,但所有已知的噬菌体都是在细菌细胞中利用细菌的核糖体、蛋白质合成时所需的各种因子、 ...
诺卡氏菌属于革兰氏阳性。有时抗酸的菌丝体内含诺卡氏菌类脂(LCN~A)和诺卡枝菌酸,细胞壁Ⅳ型。根据菌丝体发育的程度和断裂的迟早,将本属分为3群。本属菌广泛分布于自然界医学教育|网搜集整理。有的致病,引起人和动物的诺卡氏菌病。它们也是多种抗生素的产生菌。此外,在石油脱蜡、污水净化方面也起一定作用。代表种为鼻疽诺卡氏菌。诺卡氏菌
根霉的菌丝无隔膜、有分枝和假根,营养菌丝体上产生匍匐枝,匍匐枝的节间形成特有的假根,从假根处向上丛生直立、不分枝的孢囊梗,顶端膨大形成圆形的孢子囊,囊内产生孢囊孢子。医学教育|网搜集整理孢子囊内囊轴明显,球形或近球形,囊轴基部与梗相连处有囊托。根霉的孢子可以在固体培养基内保存,能长期保持生活力。根霉
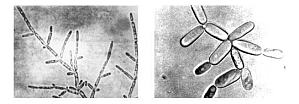
裂殖酵母属于子囊菌亚门、酵母科中的裂殖酵母亚科。细胞为椭圆形或圆柱形。无性繁殖为分裂繁殖。有时形成假菌丝。有性繁殖是营养细胞(单倍体)结合形成子囊,子囊内有1~4个或8个子囊孢子。子囊孢子是球形或卵圆形,具有酒精发酵的能力,不同化硝酸盐。八孢裂殖酵母是这一属的重要菌种。无性繁殖为裂殖,麦芽汁,25℃,培养3天,液面无菌醭,液清医学教育|网搜集整理,菌体沉于管底。在麦芽汁琼脂培养基上菌落为乳白色,无光泽,曾经从蜂蜜、粗制蔗糖 ...
螺旋菌又称幽门螺杆菌或幽门螺旋菌。 幽门螺旋菌简称HP(Helicobocton Pyloni),医学教育|网搜集整理此种菌类是由瑞典学者首先从人胃粘膜标本培养中发现的,也是近年来国内研究的热题。螺旋菌
眼虫是眼虫属生物的统称,在植物学中称裸藻医学教育|网搜集整理,是一类介于动物和植物之间的单细胞真核生物。眼虫
硝化细菌(nitrifying)是一种好氧性细菌,包括亚硝化菌和硝化菌医学教育|网搜集整理。生活在有氧的水中或砂层中,在氮循环水质净化过程中扮演着很重要的角色。 硝化细菌
球菌是一种呈球形或近似球形的细菌。根据排列方式不同,医学教育|网搜集整理可分为单球菌、双球菌、链球菌、四联球菌、八叠球菌和葡萄球菌等。 球菌
杆菌是杆状或类似杆状的细菌。广泛分布于自然界。腐生或寄生。如大肠杆菌、枯草杆菌等。各种杆菌的大小、长短、弯度、粗细差异较大。大多数杆菌中等大小长2~5um,宽0.3~1um.大的杆菌如炭疽杆菌(3~5um×1.0~1.3um),小的如野兔热杆菌(0.3~0.7um×0.2um)。菌体的形态多数呈直杆状,也有的菌体微弯。菌体两端多呈钝圆形,少数两端平齐(如炭疽杆菌),也有两端尖细(如梭杆菌)或末端膨大呈棒状(如白喉杆菌)。医学教育|网搜集整 ...

