DETECTION OF ß-GALACTOSIDASE AND ALKALINE PHOSPHATASE ACTIVITIES IN TISSUE
互联网
DETECTION OF ß-GALACTOSIDASE
The E. coli lacZ gene encoding ß-galactosidase (ß-gal) is the classical histochemical reporter gene (Beckwith, 1980). It can be detected using a variety of substrates, all of which have galactose linked through a ß-D-glycosidic linkage to a moiety whose properties change upon liberation from galactose (Wallenfels and Weil, 1972). Several substrates yield colored or fluorescent soluble products which are useful when quantifying ß-gal activity (McCaman and Robins, 1959, Miller 1972) or visualizing transduced cells live in vivo (Krasnow et al. 1991, Nirenberg and Cepko, 1993, Lin et al. 1994). The fluorescent products can even be used to kill cells in vivo (Nirenberg and Cepko, 1993). However, for localization of cells containing transduced lacZ, chromogenic substrates that yield a precipitated product are desirable (Holt and O'Sullivan, 1958, Pearse 1954, Pearson et al 1963). The most common such substrate is an indole derivative, 5-bromo-4-chloro-3-indolyl-ß-D-galactoside (X-gal, Holt and Sadler, 1958).
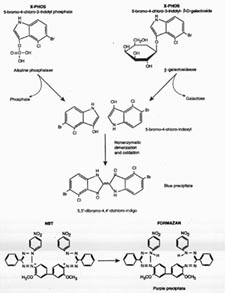
When ß-gal cleaves the glycosidic linkage in X-gal, a soluble, colorless indoxyl monomer is produced. Subsequently, 2 of the liberated indoxyl moieties form a dimer which is nonenzymatically oxidized (Figure 1) . The resultant halogenated indigo is a very stable and insoluble blue compound (Holt and Sadler, 1958). The dimerization and oxidation reactions require transfer of an electron , which is facilitated by electron acceptors of the proper redox potential (Cotson and Holt, 1958). The ferric and ferrous ions included in most X-gal reaction buffers provide this function (Lojda, 1970). Tetrazolium salts, which can serve as the final electron acceptors, also can be added, and precipitate when reduced to form colored formazan compounds (Altman, 1972). Phenazine methosulfate (PMS) can further increase this reaction rate by quantitatively reducing tetrazolium salts (Altman, 1972). Alternative staining protocols that yield different colored products have been developed based upon these considerations and will be discussed at the end.
X-GAL STAINING OF INTACT TISSUE (WHOLE MOUNTS)
- Dissect embryos or small pieces of tissue (e.g. the size of a retina from an adult rat works well with this procedure) into PBS containing 2mM MgCl2 (PBS + Mg2+) on ice.
PBS(10X)- 80 g NaCl
- 2 g KCl
- 11.5g Na2HPO4
- 2 g KH2PO4
- in 1 liter H20
- Fix in 0.5% glutaraldehyde in PBS + Mg2+ or 2.0%- 4.0% paraformaldehyde in PBS + Mg2+ on ice for 30' to several hours. The amount of time should be determined empirically. Generally, it is wise to minimize time in fixative as the lacZ encoded enzyme can be overfixed. Fixation with 0.5% glutaraldehyde gives superior staining relative to paraformaldehyde fixation, but can also preserve endogenous enzyme activity to the point where the signal from endogenous activity is confounding.
0.5% glutaraldehyde- Make from a 25.0% stock immediately before use. You can buy a 25% solution (Sigma) and freeze-thaw it many times.
4.0% paraformaldyde- 4 g paraformaldehyde
- 2 mM MgCl2 (0.2 ml of a 1 M stock)
- 1.25 mM EGTA (0.25 ml of a .5M EGTA stock, pH 8.0)
- in 100 ml PBS, pH 7.2-7.4 Heat about 80ml H2 O to 600 C and add paraformaldehyde; add 1-2 drops of 10M NaOH to get paraformaldehyde in solution. Cool to room temperature, add 10 ml 10X PBS, adjust pH with HCl, add MgCl2 and EGTA, and make up to 100ml with H2O. The solution can be stored at 40 C 1-2 weeks.
- Make from a 25.0% stock immediately before use. You can buy a 25% solution (Sigma) and freeze-thaw it many times.
- Rinse in many changes of PBS. Residual fixative can inhibit the enzyme. Rinsing overnight is fine, but waiting for several days at this step may decrease ß-gal activity, and paraformaldehyde fixed tissue can become "unfixed".
- Dilute the X-gal stock into X-gal Reaction Buffer and incubate with the tissue 2-4 hours at 370
C.
X-gal Reaction Buffer
- 35 mM potassium ferrocyanide (can vary from 5-35 mM)
- 35 mM potassium ferricyanide (can vary from 5-35 mM)
- 2 mM MgCl2
- 0.02% Nonidet P-40 (NP-40) (diluted from 10% stock solution)
- 0.01% Na deoxycholate (diluted from 10% stock solution) in PBS
X-gal reaction buffer can be stored for at least one year at room temperature in foil-covered container.
- X-gal stock(40X) 40 mg/ml X-gal in dimethylformamide
- Store at -200 C in foil covered glass container.
- Rinse many times in PBS until the solution no longer turns yellow. This usually takes about 5 changes. O/N rinse is fine.
- View under bright field optics for optimal detection.
Notes
- Although we have seen reduced activity of ß-gal after treatment with organic solvents, lyophilization followed by permeabilization in acetone has been used successfully in place of aldehyde fixation for X-gal staining in C. elegans (Fire, 1992).
- The amount of ferricyanide and ferrocyanide can be varied. A more rapid precipitation is achieved with the higher concentration, but the higher concentration can lead to a greenish precipitate (Prussian Green) in tissue upon prolonged staining.
- The ferricyanide and ferrocyanide can form a blue precipitate (Prussian blue) upon reaction with free ferric ion. Do not use metal forceps to manipulate the tissue while it is in the X-gal detection buffer.
- iv. All tissues have endogenous, lysozomal ß-gal. Its pH optimum is very low, and thus it is not very active in the pH 7.4 buffer used here. However, some tissues also have a cytosolic form of ß-gal which can show enough activity to be confounding. The variables that affect background staining include the fixative type, length of time in fixative, pH of the buffer, and amount of time in the staining solution (Rosenberg et al 1992). If after varying these parameters, background is still a problem, one can detect the lacZ enzyme by immunohistochemistry. Monoclonal and polyclonal antibodies are available (e.g. from 5 prime, 3 prime, Boehringer-Mannheim and Cappel).
- Tris buffer was tried in place of PBS in the reaction buffer, with no success.
- The detergents in the X-gal reaction buffer need not be included, but usually do not reduce staining, and for some tissues they increase staining.
- The indigo product is soluble in organic solvents and thus one should minimize exposure to such solvents after formation of the product. Nonetheless, fixation with glutaraldehyde (but not formaldehyde) allowed preservation of enough indigo to make it through the various solvents required for embedding for electron microscopy (EM) (Snyder et al. 1992).
- Double staining of cells for ß-gal with X-gal and a cellular antigen using antibodies is difficult, though possible (Snyder et al 1992, Vaysse and Goldman, 1990). The indigo product of X-gal absorbs in the wavelengths emitted by the standard fluorescent-conjugated antibodies and is so dark that it can obscure the products from either horseradish peroxidase or alkaline phosphatase-conjugated antibodies. However, by carefully controlling the reaction time in X-gal so that only a small amount of indigo is produced (Vaysse and Goldman, 1990), by having ß-gal localized to the nucleus when the cellular antigen is non-nuclear (Bonnerot et al 1987), or by using antibodies to detect both ß-gal and the cellular antigen, one can overcome these problems.
- If nitroblue tetrazolium (NBT) salt is added to the X-gal reaction in place of iron, a purple precipitate will result. This can be a faster and more sensitive reaction than X-gal alone. A stock solution of NBT can be prepared by adding 50 mg NBT per ml of 70% dimetnylformamide and stored at -20o C. Final working concentrations are 0.25 - 1.0 mg/ml. Phenazine methosulfate (PMS) can be added in conjunction with NBT to increase the reaction rate further. Prepare a 100X stock of 2 mg/ml in H2O and use immediately. PMS is very unstable.
- To aid in identification of stained cells, the tissue can be processed for cryostat sections as discussed below. In addition, staining cryostat sections may give higher signal than whole mounts so if you are not sure that the whole mount procedure gave the maximal staining, stain sections as below. Alternatively, for simply visualizing the stained cells produced during staining of whole mounts, paraffin sections can be made (paraffin embedding destroys ß-gal activity so there is no point in staining paraffin sections). Embed in paraffin using minimum necessary times for the tissue of interest as the solvents can partially dissolve indigo. For glutaraldehyde-fixed mouse retina, which is approximately 250 microns thick, the following procedure was used. Dehydrate through graded ethanols (50%, 70%, 95%, 100%, 100%) for 20 min each. Clear in xylene, 2 x 15 min. Infiltrate with 1:1 mix of xylene and paraffin, 65O C for 30 min. Paraffin, 2 x 15 min. Embed in paraffin.
X-GAL STAINING OF TISSUE SECTIONS
The procedure for staining tissue sections is very similar to the protocol for intact tissue. Section staining should be used when a whole mount cannot be used due to the size of the tissue.
- Fix tissue in fixatives listed above using perfusion if possible and follow with immersion in fixative at 4O
C for several hours. Rinse briefly in PBS, then sink in 30% sucrose in PBS + Mg2+at 4O
C.
Fixation times will vary with the size and nature of the tissue. For embryonic chick brains of E10 or older and for postnatal rat or mouse brains we incubate up to 8 hours in fixative. Perfusion may not be necessary for all tissues and shorter fixation times may be preferable as X-gal staining may be decreased by lengthy fixation.
- Embed tissue in OCT (Miles) or gelatin/sucrose mounting medium and freeze on metal chucks cooled in liquid N2.
Gelatin/sucrose embedding gives better frozen sections for embryonic tissue than does OCT.
- Gelatin/sucrose embedding solution
- 7.5% gelatin (porcine skin, Sigma)
- 15% sucrose
- 0.05% sodium azide
- in 1X PBS
- Cut cryostat sections and mount on silane-coated (Rentrop et al., 1986) or gelatin-coated slides; air-dry O/N. Sections up to 90 m m thick (the thickest we have tried) have been successfully stained.
- Gelatin Solution for subbing slides
- 2 g gelatin
- 0.1 g chromium potassium sulfate (chrome alum)
- in 200 ml H20
Heat H20 to 60oC. Dissolve chrome alum, then gradually dissolve gelatin. Filter before use. Can increase or decrease the percentage of gelatin. Load slides in racks, dip quickly, and air-dry overnight.
- Fix sections to slides in 4% paraformaldehyde for 10-15 minutes at 4O C.
- Rinse slides in PBS + Mg2+ twice, for 10 minutes each, at 4O C.
- Stain slides in X-gal Reaction Buffer for 1-24 hours at 37O C.
- Rinse slides in PBS 3 times, for 10 minutes each, or until solution is no longer yellow. Slides can be left in PBS O/N.
- Coverslip in gelvatol.
GELVATOL PREPARATION
We have revised the protocol published by Rodriguez and Deinhardt (1960).
- Take 200 ml of 0.01M KH2PO4 (about pH 5.0) and add enough 0.01 M Na2HPO4 to bring the pH up to 7.2.
- Take 250 ml of the 0.01M KH2PO4/Na2HPO4 and add 2.05 g NaCL to give a 0.14M NaCL concentration.
- Dissolve 62.5 g Gelvatol (Air Products) in the 250 ml of 0.01 M KH2PO4/Na2HPO4/.14 M NaCL. Stir on magnetic stirrer in warm room for a few hours.
- Add glycerol in an amount equal to one-half the total volume of the Gelvatol buffered saline solution and stir overnight at room temperature.
- Centrifuge the Gelvatol solution at 12,000 rpm for 15 min. in 30 ml Corex tubes in Beckman J2-21 centrifuge at room temperature to remove undissolved particles.
- Pipette the supernatant into small screw cap bottles. Check pH of Gelvatol solution. It should be between pH 6 and 7.
- Store Gelvatol solution at 4 o C. Screw caps on tightly to prevent evaporation. Do not leave Gelvatol uncapped for longer than necessary when working with it.
Notes
- The amount of time for the X-gal reaction will vary according to the concentration of lacZ and the amount of endogenous ß-gal.
- If cultured cells are to be stained, fix in 0.5% glutaraldehyde in PBS or in 4.0% paraformaldehyde in PBS for 5' at room temperature and proceed from step 5 above. Fixation for > 5 minutes can lead to decreased enzyme activity.
STAINING FOR ALKALINE PHOSPHATASE ACTIVITY
Phosphatase genes are useful reporter genes as several histochemical methods that yield precipitated, highly colored and/or electron dense products have been devised. A chromogenic substrate, 5-bromo-4-chloro-3-indolyl phosphate (X-Phos), which is very similar to X-gal, can be employed for detection and leads to production of a blue precipitate (Figure 1). In addition, the tetrazolium salt NBT, (Altman, 1972) can be used in conjunction with X-Phos as the final electron acceptor for the indoxyl dimerization reaction. When reduced, NBT forms a purple precipitate.
Of the cloned phosphatases, the human placental alkaline phosphatase gene, PLAP (Kam et al 1985), is perhaps the most useful as it is very heat stable and is resistant to some chemical inhibitors that are active on other endogenous alkaline phosphatases. PLAP has been used as a histochemical reporter fairly recently (Berger et al 1987, Henthorn et al 1988, Fields-Berry et al 1992). It has not been used as widely as lacZ and thus its neutrality needs to be established. LacZ and PLAP have different strengths and weaknesses regarding their distribution and histological detection. PLAP is normally associated with membranes and thus PLAP activity and defines the outer surface of transduced cells, including neuronal processes. We have found PLAP staining in retinal ganglion cell axons several centimeters from cell bodies (Fekete and Cepko, unpublished). However, there are times when neuronal cell bodies are not well defined, which can make it difficult to count cells (Halliday and Cepko, 1992 and Fekete et al, 1994). ß-gal activity visualized by X-gal typically does not fill cells, particularly long processes, but often gives good staining of cell bodies and thus it is easy to count cells. Frequently, X-gal staining is fairly perinuclear and sometimes it is punctate as well. When X-gal precipitate is viewed under the EM, it appears that it localizes to the nuclear envelope and the golgi and endoplasmic reticulum (Snyder et al 1992). For PLAP, there exists a procedure in which lead is precipitated very close to the location of the enzyme (Hugon and Borgers, 1966). The lead precipitate is superior to that of X-gal and thus PLAP is the enzyme of choice when electron microscopy is required.
There are endogenous alkaline phosphatase activities that may lead to difficulties in detection of transduced PLAP (McComb et al., 1979). In most cases, background activities can be minimized with heat and chemical inhibitors that leave PLAP active (Zoellner and Hunter, 1989). In addition, monoclonal and polyclonal antibodies specific to PLAP can be purchased (Dako, Zymed, Medix, and Accurate Chemical and Scientific Co.) when the background is not surmountable using the various inhibitors.
X-PHOS/NBT STAINING OF WHOLE MOUNTS
See above protocol for X-gal staining of whole mounts and perform steps 1 and 2. 3. Heat tissue for at least 30' at 650 C.For staining of embryonic chick diencephalon (one of the areas of the brain with the highest background), this step was increased to 1.5 hours. It may be that even longer heat treatment could benefit specific staining in areas with high background.
4. Incubate tissue in X-Phos/NBT Detection Buffer for 15' at room temperature.
XPhos/NBT Detection Buffer (Buffer 3 as described for Genius kit by Boehringer-Mannheim)
- 100 mM Tris-HCl, pH 9.5
- 100 mM NaCl
- 50 mM MgCl2
Store at room temperature. Tends to precipitate over several weeks. This does not seem to noticeably affect the staining.
5. Incubate in X-Phos/NBT Reaction Solution for 1 to several hours at room temperature. Cover the reaction with foil to reduce background.
- X-Phos/NBT Reaction Solution
- 50 ul of 100X X-Phos stock
- 100 ul 50X NBT stock
- 50 ul of 100X levamisole stock, optional
- in 5 ml X-Phos Detection Buffer
- Mix X-Phos, NBT, and levamisole with the Detection Buffer immediately before using.
Stocks
- X-Phos (100X):10mg/ml 5-bromo-4-chloro-3-indolyl-phosphate (also referred to as BCIP) in H20. Store in the dark as aliquots at -200 C. Can be frozen and thawed several times.
- NBT (50X):50 mg/ml nitro blue tetrazolium in 70% dimethylformamide, 30% H2O. Store at -200 C in a glass container covered with foil. Does not freeze at this temperature.
- Levamisole (100X): 50 mM in H20. Store at -200 C.
- PMS (100X): 2 mg/ml phenazine methosulfate in H20. Use immediately.
6. Rinse in 20mM EDTA in PBS for 2-4 hours.
Tissue can be stored in the dark at 40 C in PBS + EDTA or 30% sucrose in PBS + EDTA + 0.05% sodium azide for many months, although the background clearly increases over time. Tissue can then be sectioned as above for X-gal stained whole mounts.
Notes
- 0.5% glutaraldehyde decreased PLAP activity in embryonic chick brain cells stained in whole mounts, but not in chicken embryo fibroblasts cultured in vitro. Fixation of chick brain whole mounts thus is typically done in 4% paraformaldehyde in PBS for 2-4 hours at 4O C. In areas where background alkaline phosphatase activity is a problem, increasing the time in 4% paraformaldehyde, even up to several days, can decrease endogenous background without significantly decreasing PLAP activity.
- Chick retinas and cerebella have been kept in PBS at 4O C for at least one month after fixation and rinsing in PBS with no appreciable loss of signal in X-Phos staining.
- Background staining can be due to endogenous alkaline phosphatase activities or reduction of NBT from other sources (e.g. NADPH). It is also enhanced by light. The most effective inhibitor is heat. To reduce background further, one or more of the following inhibitors can be tried in addition to heat (they are added to the reaction mix): 0.5 mM levamisole (L[-]-2,3,5,6-tetrahydro-6-phenylimidazo{2,1-b}thiazole), 2 mM mercuric chloride, 5 mM L-leucyl-glycyl-glycine, 1 mM EDTA, 1 mM L-phenylalanine-glycyl-glycine, 0.2 M lysine HCl or 0.3 mM sodium arsenate (Zoellner and Hunter, 1989). We found that levamisole was the second most useful treatment (after heat) in reducing the background staining in brains, although it also reduced PLAP staining slightly in some cases.
- If background staining is not a problem, the reaction can be continued for up to 2 days.
X-PHOS/NBT STAINING OF SECTIONS
Cryostat sections can be stained for PLAP activity. Follow the protocol listed above for X-gal through step 5.
6.Transfer slides to preheated PBS at 65O C and heat for 30 minutes.
7. Rinse slides in room temperature PBS for 5 minutes.
8. Rinse slides in X-Phos/NBT Detection Buffer for 10 minutes.
9. Stain slides in X-Phos/NBT Reaction Mix for 1 to 12 hours at room temperature. Cover with foil during and after staining. The time in the reaction buffer will depend on the level of PLAP expression and endogenous background. If background is low, staining can continue for 2 days.
10. Rinse slides in PBS + 20 mM EDTA, 3x 10 minutes.
11. Mount in Gelvatol (+ 20 mM EDTA if desired).
Storing slides at -80o C helped prevent background staining from increasing.
Notes
- If cultured cells are to be stained, fix in 0.5% glutaraldehyde in PBS or in 4.0% paraformaldehyde in PBS for 5' at room temperature and proceed from step 6 above.
- Processing cultured cells grown on glass through the procedure for paraffin embedding prior to staining preserved enough PLAP activity that positive cells were visible. If paraffin sections are desirable, it is worth trying pilot experiments using the tissue of interest, varying the fixation, and minimizing the times in organic solvents.
X-gal AND X-PHOS/NBT STAINING
ß-gal and PLAP can be detected in the same tissue and even in the same cell. In order to process tissue for both activities, staining with X-gal must be done first as the lacZ-encoded enzyme is destroyed by the heating step used to reduce endogenous alkaline phosphatase activities. To combine the protocols, proceed through the X-gal reaction as described above for either whole mounts or sections. You may wish to stop and examine the results before moving to PLAP staining. The indigo product of X-gal can be easier to detect prior to carrying out the X-Phos reaction as background alkaline phosphatase staining can obscure it, particularly in whole mounts. Rinse the tissue very well with PBS prior to PLAP staining as residual ß-gal activity in the presence of X-gal and NBT can enable ß-gal+, PLAP- cells to turn purple. Follow the above protocols for PLAP staining.
A COMPARISON OF ALTERNATIVE SUBSTRATES FOR ß-gal AND PLAP
As described above, there are a number of alternative substrates for both enzymes. The following discussion concerns these substrates and summarizes our experiences with them relative to the those described above.
ß-gal SUBSTRATES:
X-gal is a good choice as a precipitable substrate with iron as an electron acceptor. It gives a bright blue color, although the reaction is quite slow, even at 37O C. NBT added to X-gal gives an intense purple color which develops much more rapidly. PMS will increase this reaction rate even further. Tetrazolium red makes the product look greenish-blue, but increases background, making the staining more equivocal. Other modified indolyl-based compounds can be used if alternative colored precipitates are desired. Salmon-gal (Biosynth International) results in a light orange/pink precipitate which develops slowly. Adding tetrazolium salts tends to increase the background. Magenta-gal (Biosynth International) gives a light pinkish purple precipitate. Green-gal (Biosynth International) did not give discernible precipitates in our hands, except in the presence of NBT, which gave a purple precipitate.
PLAP substrates
X-Phos in the absence of NBT gives a bright blue precipitate which develops slowly over time. As a result, there may be some diffusion away from the site of enzyme activity. Iron does not work as an electron acceptor for the indolyl-based compounds at high pH, and results in large amounts of floating precipitate. Adding NBT results in a deep purple precipitate which develops more quickly. Adding PMS increases the rate of the reaction (in the presence of NBT) without adding background by quantitatively reducing the tetrazolium salt. Other modified indolyl-based compounds can be used if alternative colored precipitates are desired. Magenta-phos (Biosynth International) gives a pinkish-purple precipitate which develops at intermediate rates. The color can be distinguished from the purple color derived from X-Phos + NBT. Adding tetrazolium red or blue makes the product look more purple (less pink). Salmon-phos (Biosynth International) results in a light orange-pink precipitate which develops slowly. Adding tetrazolium red or blue gives a purple precipitate similar to XP + NBT. The Vector Red substrate kit (Vector Laboratories) results in staining that is highly variable between experiments; the precipitate can be intensely red but is usually light pink. The substrate is unstable, and begins to form floating precipitates after 20-30 minutes. Addition of tetrazolium salts is not helpful, and PMS acts as an inhibitor in this system. The substrate affords high background levels to intact tissues and thus is not useful for staining whole mounts. The blue substrate kit (Vector Laboratories) affords a very pretty peacock blue precipitate, but has the same problems as the red substrate kit, i.e. unstable substrate and high background to intact tissues. Staining is reproducible and intense, however. Adding tetrazolium salts such as NBT increases the background and results in copious amounts of non-localized precipitate.
Legend for Figure 1. Production of indigo from X-gal or X-Phos.
A. Production of the stable blue precipitate from substituted indoles proceeds as shown. Two molecules of either X-Phos or X-gal generate 2 molecules of indoxyl which then form the indicated dimer. The "X" moiety (5-bromo-4-chloro-indolyl) in both X-gal and X-Phos is the same and thus once cleavage by the respective enzyme has occurred, the dimerization and oxidation reactions lead to the same halogenated indigo compound.
B. Nitroblue tetrazolium (NBT) is an example of a tetrazolium salt. These are a rather unstable class of compounds which precipitate when reduced to form highly colored compounds. In this case, NBT is reduced by the hydride produced by dimerization of the two indoxyls that result from cleavage of either X-gal or X-Phos (shown in (A) above). When reduced, NBT forms formazan, a dark purple precipitate.
REFERENCES
- Berger, J., A.D. Howard, L. Gerber, B.R. Cullen, and S. Udenfriend. 1987. Expression of active, membrane-bound human placental alkaline phosphatase by transfected simian cells. Proc. Natl. Acad. Sci. USA 84 :4885.
- Beckwith, J.R. 1980. Lac : The genetic system. In The Operon (ed. J.H. Miller and W.S. Reznikoff). Cold Spring Harbor Laboratory, Cold Spring Harbor, New York.
- Bonnerot, C., Rocancourt, D., Briand, P., Grimber, G., and Nicolas, J.-F. 1987. A ß-galactosidase hybrid protein targeted to nuclei as a marker for developmental studies. Proc. Natl. Acad. Sci. USA 84 :6795.
- Cotson, S. and S.J. Holt. 1958. Studies in enzyme cytochemistry IV. Kinetics of aerial oxidation of indoxyl and some of its halogen derivatives. Proc. Royal Soc. (London) 148B :506.
- Fekete, D.M., J. Perez-Miguelsanz, E.F. Ryder, and C.L. Cepko. Clonal analysis in the chicken retina reveals tangential dispersion of clonally related cells. Dev. Biol. In press, 1994.
-
- Halliday, A.L., and C. Cepko. 1992. Generation and migration of cells in the developing striatum. Neuron 9 :15.
- Henthorn, P., P. Zervos, M. Raducha, H. Harris, and T. Kadesch. 1988. Expression of a human placental alkaline phosphatase gene in transfected cells: Use as a reporter for studies of gene expression. Proc. Natl. Acad. Sci. USA 85 :6342.
- Holt, S.J., and D.G. O'Sullivan. 1958. Studies in enzyme cytochemistry I. Principles of cytochemical staining methods. Proc. Royal Soc. (London) 148B :465.
- Holt, S.J., and P.W. Sadler. 1958. Studies in enzyme cytochemistry III. Relationships between solubility, molecular association and structure in indigoid dyes. Proc. Royal Soc. (London) 148B :495.
- Hugon, J., and M. Borgers. 1966. A direct lead method for the electron microscopic visualization of alkaline phosphatase activity. J. Histochem. and Cytochem . 14 :429.
- Kam, W., E. Clauser, Y.S. Kim, Y.W. Kan, and W.J. Rutter. 1985. Cloning, sequencing, and chromosomal localization of human term placental alkaline phosphatase cDNA. Proc. Natl. Acad. Sci. USA 82 :8715.
- Krasnow, M.A., S. Cumberledge, G. Manning, L.A. Herzenberg, and G.P. Nolan. 1991. Whole animal cell sorting of Drosophila embryos. Science 251 :81.
- Lin, S., S. Yang, and N. Hopkins. 1994. LacZ expression in germline transgenic zebrafish can be detected in living embryos. Dev. Biol. 161 :77.
- Lojda, Z. 1970. Indigogenic methods for glycosidases. Histochemie 22 :347.
- McCaman, R.E., and E. Robins. 1959. Quantitative biochemical studies of Wallerian degeneration in the peripheral and central nervous systems-II. J. Neurochemistry 5 :32.
- McComb, R.B., G.N. Bowers, Jr., and S. Posen. 1979. Alkaline Phosphatase. Plenum Press, New York.
- Miller, J. 1972. Experiments in molecular genetics. Cold Spring Harbor Laboratory, Cold Spring Harbor, New York.
- Nirenberg, S., and C. Cepko. 1993. Targeted ablation of diverse cell classes in the nervous system in vivo. J. Neuroscience 13 :3238.
- Pearse, A.G. 1954. Azo dye methods in enzyme histochemistry. In International Review of Cytology vol. III (ed. G.H. Bourne, and J.F. Danielli) Academic Press, New York.
- Pearson, B., P.L. Wolf, and J. Vazquez. 1963. A comparative study of a series of new indolyl compounds to localize ß-galactosidase in tissues. Lab Investig . 12 :1249.
- Rentrop, M., B. Knapp, H. Winter, and J. Schweizer. 1986. Aminoalkylsilane-treated glass slides as support for in situ hybridization of keratin cDNAs to frozen tissue sections under varying fixation and pre-treatment conditions.Histochem . J. 18 :271.
- Rodriguez, J., and F. Deinhardt. 1960. Preparation of a semipermanent mounting medium for fluorescent antibody studies. Virology 12 :316.
- Rosenberg, W.S., X.O. Breakefield, C. DeAntonio, and O. Isacson. 1992. Authentic and artifactual detection of the E. coli laxZ gene product in the rat brain by histochemical methods. Brain Research 16 :311.
- Snyder, E.Y., D.L. Deitcher, C. Walsh, S. Arnold-Aldea, E.A. Hartwieg, and C.L. Cepko. 1992. Multipotent neural cell lines can engraft and participate in development of mouse cerebellum. Cell 68 :33.
- Vaysse, P.J.-J., and J.E Goldman. 1990. A clonal analysis of glial lineages in neonatal forebrain development in vitro. Neuron 5 :227.
- Wallenfels, K., and R. Weil. 1972. In The Enzymes . (ed. P.D. Boyer). 3rd edition. 7 :617. Academic Press, New York.
- Zoellner, H.F., and Hunter, N. 1989. Histochemical identification of the vascular endothelial isoenzyme of alkaline phosphatase. J. Histochem. and Cytochem. 37(12) : 1893.