类病毒
互联网
一、类病毒�
20 世纪 70 年代初期,美国学者 Diener 及其同事在研究马铃薯纺锤块茎病病原时,观察到病原无病毒颗粒和抗原性、对酚等有机溶剂不敏感、耐热 (70 ℃ ~75 ℃ ) 、对高速离心稳定 ( 说明其低分子量 ) 、对 RNA 酶敏感等特点。所有这些特点表明病原并不是病毒,而是一种游离的小分子 RNA 。从而提出了一个新的概念―类病毒 (Viroid) 。在这个概念提出之前,人们一直认为,由蛋白质和核酸两种生物多聚体构成的体系,是原始的生命体系,从未怀疑病毒是复杂生命体系的最低极限。�
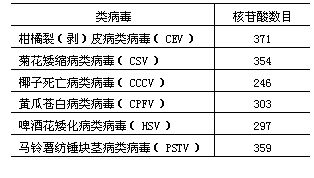
类病毒是一类能感染某些植物致病的单链闭合环状的 RNA 分子。类病毒基因组小,分子量为 1 × 10 5 。目前已测序的类病毒变异株有 100 多个,其 RNA 分子呈棒状结构,由一些碱基配对的双链区和不配对的单链环状区相间排列而成。它们一个共同特点就是在二级结构分子中央处有一段保守区。类病毒通常 246~399 个核苷酸。如马铃薯纺锤块茎类病毒 ( Potato spindle tuber viroid , PSTVd , Vd 是用来与病毒加以区别 ) 是由 359 个核苷酸单位组成的一个共价闭合环状 RNA 分子,长约 50A~70nm( 见图 3 ・ 16) 。 �
所有的类病毒 RNA 没有 mRNA 活性,不编码任何多肽,它的复制是借助寄主的 RNA 聚合酶 II 的催化,在细胞核中进行的 RNA 到 RNA 的直接转录。�
图 3.16 类病毒的结构图
类病毒能独立引起感染,在自然界中存在着毒力不同的类病毒的株系。 PSTVd 的弱毒株系只减产 10% 左右,而强毒株可减产 70%~80% 。�
所有的类病毒均能通过机械损伤的途径来传播,经耕作工具接触的机械传播是在自然界中传播这种病害的主要途径。有的类病毒,如 PSTVd 还可经种子和花粉直接传播。类病毒病与病毒病在症状上没有明显的区别,病毒病大多数典型症状也可以由类病毒引起。类病毒感染后有较长的潜伏期,并呈持续性感染。�
不同的类病毒具有不同的宿主范围。如对 PSTVd 敏感的寄主植物就数以百计,除茄科外,还有紫草科、桔梗科、石竹科、菊科等。柑桔裂皮类病毒 ( Citrus exocortis viroid , CEVd) 的寄主范围比 PSTVd 要窄些,但也可侵染蜜柑科,菊科,茄科,葫芦科等 50 种植物。�
类病毒的发现,是 20 世纪下半叶生物学上的重要事件,开阔了病毒学的视野。它为进一步研究植物中可能存在的类病毒病开辟了一个新的方向。�
二、朊病毒�
美国学者 S. B. Prusiner 因发现了羊瘙痒病致病因子―朊病毒( 1982 年),而获得了 1997 年的诺贝尔生理和医学奖。朊病毒 (Virino) 亦称蛋白侵染因子 (Prion, Proteinaceous infectious agents) ,是一种比病毒小、仅含有疏水的具有侵染性的蛋白质分子。
纯化的感染因子称为朊病毒蛋白 ( Prion protein , PrP) 。致病性朊病毒用 PrP SC 表示,它具有抗蛋白酶 K 水解的能力,可特异地出现在被感染的脑组织中,呈淀粉样形式存在。
许多致命的哺乳动物中枢神经系统机能退化症均与朊病毒有关 ,如人的 库鲁病( kuru ,一种震颤病)、克雅氏症 (Creutzfeldt-Jakob Disease, CJD ,一种早老年痴呆病 ) 、致死性家族失眠症 (Fatal Familiar Insomnia, FFI) 和动物的羊瘙痒病 (Scrapie) 、牛海绵状脑病 (Bovine Spongiform Encephalopathy , BSE 或称疯牛病 mad cow disease) 、猫海绵状脑病 (Feline Spongifoem Encephalopathy , FSE) 等。
正常的人和动物细胞 DNA 中有编码 PrP 的基因,其表达产物用 PrP c 表示,相对分子量为 33~35kDa 。正常细胞表达的 PrP c 与羊瘙痒病的 PrP sc 为同分异构体, PrP C 与 PrP SC 有相同的氨基酸序列, PrP C 有 43% 的α螺旋和 3% 的β折叠,而 PrP SC 约有 34% 的α螺旋和 43% 的β折叠。多个折叠使 PrP SC 溶解度降低、对蛋白酶的抗性增加。
既然 PrP SC 是一种蛋白质而且不含任何核酸,那么它在人或动物体内又是如何进行复制,如何进行传播的呢 ? Prusiner 等提出了杂二聚机制假说,既 PrP SC 单分子为感染物,从 PrP C 单体分子慢慢改变构象,形成 PrP SC 单体分子,中间经过 PrP C - PrP SC 杂二聚物,然后再转变为 PrP SC -PrP C 。在这个过程中,有未知蛋白( protein X )可能起着调整 PrP C 转化或维持 PrP SC 形态的作用。这个二聚物解离又释放新的 PrP SC ,因此不断“复制”下去(图 3.17 )。
图 3.17 PrP C 与 PrP SC 的分子结构模式图 PrP C :正常型蛋白 PrP SC :致病型蛋白
图 3.18 朊蛋白的构型转换
朊病毒的发现在生物学界引起震惊,因为它与目前公认的“中心法则”即生物遗传信息流的方向是“ DNA ? RNA →蛋白质”的传统观念相抵触。 Pursiner 等人阐明羊瘙痒病的发病机制是由于朊病毒分子构象的改变而致病。这一发现开辟了病因学的一个新领域,可能对其他传染性海绵状脑病的发病原理和病因性质,提供一条新的思路,对生物科学的发展具有重大意义。
三、拟病毒和卫星 RNA
20 世纪 80 年代以来,在澳大利亚陆续从绒毛烟、苜蓿、茛菪和地下三叶草上发现了四种新的植物病毒。这些病毒的蛋白质衣壳内都含有两种 RNA 分子,一种分子量为 1.5 × 10 6 Da 的线状 RNA 1 ,另一种为分子量约为 10 5 Da 的类似于类病毒的环状 RNA 2 ,这种 RNA 2 分子被称为拟病毒 (Virusoid) 。拟病毒有两种分子结构,一是环状 RNA 2 ,二是线状 RNA 3 。 RNA 2 和 RNA 3 是由同一种 RNA 分子所呈现的两种不同构型,其中 RNA 3 可能是 RNA 2 的前体,即 RNA 2 是通过 RNA 3 环化而形成的。拟病毒在核苷酸组成、大小和二级结构上均与类病毒相似,而在生物学性质上却与卫星 RNA(satellite RNA) 相同,如: ① 单独没有侵染性,必需依赖于辅助病毒才能进行侵染和复制,其复制需要辅助病毒编码的 RNA 依赖性 RNA 聚合酶。 ② 其 RNA 不具有编码能力,需要利用辅助病毒的外壳蛋白,并与辅助病毒基因组 RNA 一起包裹在同一病毒粒子内。 ③ 卫星 RNA 和拟病毒均可干扰辅助病毒的复制。 ④ 卫星 RNA 和拟病毒同辅助病毒基因组 RNA 比较,它们之间没有序列同源性。根据卫星 RNA 和拟病毒的这些共同特性,现在也有许多学者将它们统称为卫星 RNA 或卫星病毒。